Oerkiemcel



Oerkiemcellen, primordiale kiemcellen (engels primordial germ cells (PGC)) of oergeslachtscellen zijn de voorwaarde voor geslachtelijke voortplanting. Bij het menselijke embryo ontstaan ze in de derde week na bevruchting in de wand van de dooierzak. In de zesde week migreren ze naar het stelsel van geslachtsklieren. Bij alle gewervelde dieren worden de oerkiemcellen al vroeg in de ontwikkeling gescheiden van de lichaamscellen (Somatische cellen). Kiemcellen komen voort uit een oerkiemcel.[1]
Belangrijke stappen
[bewerken | brontekst bewerken]- Migratie van de oerkiemcellen naar de zich ontwikkelende geslachtsklieren
- Mitotische proliferatiefase
- Doorlopen van de meiose
- Differentiatie naar rijpe gameten
Migratie
[bewerken | brontekst bewerken]Oerkiemcellen behoren tot de eerste cellijnen die zich tijdens de ontwikkeling vormen[2] en zijn de voorlopers van gameten.[3] Er wordt gedacht dat het proces van oerkiemcelmigratie tijdens de evolutie geconserveerd is gebleven en niet de specifieke mechanismen daarin, aangezien de beweging van de oerkiemcellen op een chemische stimulus lijkt te zijn geleend van bloedcellen, zenuwcellen en het mesoderm.[2] Bij de meeste organismen begint de oerkiemcelmigratie aan de achterkant van het embryo.
Dit proces verschilt in de meeste gevallen van oerkiemcelproliferatie met uitzondering van zoogdieren, waarbij beide processen tegelijkertijd plaatsvinden. Bij de meeste zoogdieren vindt eerst specificatie plaats, gevolgd door migratie, waarna het proliferatieproces in de geslachtsklieren begint. Oerkiemcellen hebben interactie met een breed scala aan celtypen terwijl ze van de epiblast naar de geslachtsklieren gaan.[2] De oerkiemcellen migreren door amoeboïde bewegingen naar de ontwikkelende geslachtsorganen, waar zij bij het begin van de 5de week aankomen en zich erin integreren. Het traject dat de oerkiemcellen afleggen is bij bepaalde dieren zeer complex met onderliggende somatische cellen, overschrijden epitheelbarrières en reageren op signalen uit hun omgeving tijdens de actieve migratie.[4] Bij veel soorten moet een epitheel worden gekruist tijdens de migratie van de oerkiemcellen. Veranderingen in de celadhesie werden waargenomen in oerkiemcellen tijdens het verlaten van het endoderm en tijdens de initiatie van de actieve migratie.[4] Actieve migratie vindt plaats terwijl de oerkiemcellen zich verplaatsen naar de zich ontwikkelende somatische geslachtsklieren.[4] Effectieve migratie vereist een langwerpige cel en polariteit.[2] Er zijn signalen uit de omgeving nodig om de mobiliteit van de oerkiemcellen te initiëren en in stand te houden.[4] De motiliteit van de oerkiemcellen wordt geactiveerd door specifieke moleculaire reactiepaden.[3]
Oerkiemcelmigratie bij ongewervelde dieren
[bewerken | brontekst bewerken]Bij de bananenvlieg duurt het hele migratieproces naar schatting tien uur.[5] Het begint met de vorming van oerkiemcellen, doordat delende kernen omringd raken door celmembranen. Ze komen voor bij de achterste pool van het embryo.[6] Deling van de kernen stopt op het moment dat ze een celmembraan gevormd hebben.[4] Er wordt ook aangenomen dat het transcriptieproces van oerkiemcellen actief wordt onderdrukt zodra ze zijn gevormd.[4]
Bij bananenvlieg begint de oerkiemcelmigratie met een passieve beweging langs de dorsale zijde van het embryo, tijdens gastrulatie.[5] Dit wordt gevolgd door een meer passieve beweging, als gevolg van de invaginatie van het achterste middendarmprimordium, wat leidt naar de oerkiemcellen in het midden van het embryo, omgeven door epitheelcellen die naar zichzelf zijn teruggevouwen.[5] Er vindt dan een splitsing plaats in twee groepen, respectievelijk links en rechts, terwijl ze actief lateraal door het epitheel migreren om de darmen te verlaten, mogelijk gemaakt door signalering van de fibroblastgroeifactor (FGF) en een op afstoting gebaseerd mechanisme dat gebruik maakt van enzymen die worden gecodeerd door het Wunen-gen.[4][5][7] Dit wordt gevolgd door een actieve beweging dorsaal langs de basale zijde van het embryo.[5] Door gerichte migratie - waarvoor meerdere genen nodig zijn, waaronder het Columbus (clb)-gen, dat codeert voor bananenvlieg HMG-CoA-reductase - bewegen de oerkiemcellen zich naar de somatische gonadale voorlopercellen en verenigen zich ermee.[4][7] Deze twee verenigde celtypen migreren vervolgens samen naar voren, totdat ze versmelten in de embryonale geslachtsklier op de toekomstige plaats van de volwassen geslachtsklier.[5]

Oerkiemcelmigrati bij gewervelde dieren
[bewerken | brontekst bewerken]Bij de ontwikkeling van gewervelde dieren verschilt de locatie waar oerkiemcellen worden gespecificeerd en de daaropvolgende migratieroutes die ze volgen per soort.[2]

Kippen
[bewerken | brontekst bewerken]Oerkiemcellen van kippen worden aanvankelijk gespecificeerd in de pellucida (een eencellige dikke, doorschijnende epiblastlaag die boven de subkiemruimte ligt).[2][8] Na de vorming van de primitieve streep gaan de oerkiemcellen naar het kiemhalveboogvormige gebied van de embryonale schijf.[2] In tegenstelling tot de meeste modelorganismen waarbij de migratie van oerkiemcellen voornamelijk via het darmepitheel plaatsvindt, migreren de oerkiemcellen van kippen door het embryonale vasculaire epitheel.[4] Zodra ze de haarvaten hebben verlaten, vindt de laatste fase van de migratie plaats langs het dorsale mesenterium naar de zich ontwikkelende geslachtsklieren.[2]
Muizen
[bewerken | brontekst bewerken]Bij muizen worden oerkiemcellen gespecificeerd in de proximale epiblast en migreren vervolgens door de primitieve streep naar het endoderm.[4] De oerkiemcellen nestelen zich vervolgens in het epitheel van de achterdarm en zullen van daaruit via het dorsale mesenterium naar het mesoderm migreren.[2][4] Er is dan sprake van bilaterale migratie van de oerkiemcellen naar de zich ontwikkelende gonadale richels, die een patroon volgt dat sterk lijkt op dat van de bananenvlieg.[2]
Zebravis
[bewerken | brontekst bewerken]Zebravisoercellen worden gespecificeerd op vier verschillende locaties in het vroege embryo via overerving van kiemplasma (een mengsel van RNA en eiwit dat vaak wordt geassocieerd met mitochondriën).[9][4] Oerkiemcellen van deze vier locaties zullen vervolgens dorsaal migreren na downregulatie van het rgs14a G-proteïne dat E-cadherine reguleert.[2] Downregulatie zal resulteren in een verminderde cel-celadhesie, waardoor de oerkiemcellen zich kunnen losmaken en het migratieproces kan beginnen. De migratie van de oerkiemcellen gaat vervolgens verder richting de zich ontwikkelende somieten 1-3.[10] Deze beweging wordt gecoördineerd door de expressie van de chemo-attractant SDF1A (stromale cel-afgeleide factor 1a).[4] De uiteindelijke migratie naar de zich ontwikkelende gonaden vindt 13 uur na de bevruchting plaats, waarna de oerkiemcellen samenvloeien met de somatische gonadevoorlopercellen.[4] Het hele proces duurt ongeveer 24 uur.[4]
Gametogenese met mitose en meiose
[bewerken | brontekst bewerken]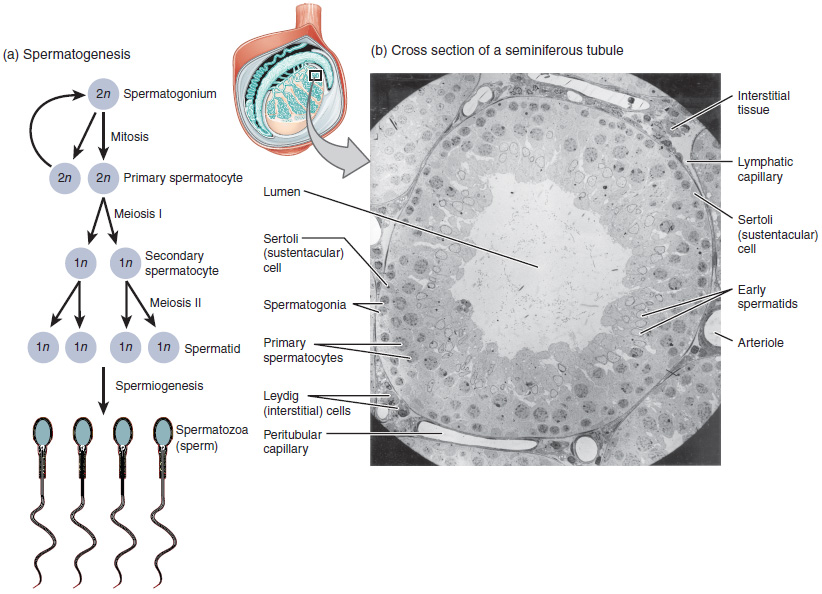
Bij de gametogenese worden mannelijke en vrouwelijke gameten gevormd door mitose en meiose. Gametogenese wordt onderscheiden in:
- spermatogenese, de vorming van zaadcellen,
- oögenese of ovogenese, de vorming van eicellen.
Gonocyten zijn de voorlopers van de spermatogonia die zich in de teelballen differentiëren uit oerkiemcellen rond week 7 van de embryonale ontwikkeling en blijven bestaan tot de postnatale periode, wanneer ze spermatogonia worden.
Zie ook
[bewerken | brontekst bewerken]- Gonocyt
- Primordial germ cell migration
Literatuur
[bewerken | brontekst bewerken]- Heinrich Zankl: Von der Keimzelle zum Individuum. Verlag C. H. Beck, 2001, ISBN 3-406-44749-X.
- Thomas W. Sadler: Medizinische Embryologie. Verlag Thieme, 2003, ISBN 3-13-446610-4.
- ↑ Dietrich Starck: Embryologie. Ein Lehrbuch auf allgemein biologischer Grundlage. 3. Auflage. Thieme, Stuttgart 1975, S. 1–45: Keimzellen und Keimzellenbildung.
- ↑ a b c d e f g h i j k (en) Cantú, Andrea V., Laird, Diana J. (1 oktober 2017). A pilgrim's progress: Seeking meaning in primordial germ cell migration. Stem Cell Research 24: 181–187. ISSN: 1873-5061. PMID 28754603. PMC 5634928. DOI: 10.1016/j.scr.2017.07.017.
- ↑ a b (en) Grimaldi, Cecilia, Raz, Erez (1 april 2020). Germ cell migration—Evolutionary issues and current understanding. Seminars in Cell & Developmental Biology 100: 152–159. ISSN: 1084-9521. PMID 31864795. DOI: 10.1016/j.semcdb.2019.11.015.
- ↑ a b c d e f g h i j k l m n o (en) Barton, Lacy J., Leblanc, Michelle G., Lehmann, Ruth (1 oktober 2016). Finding their way: themes in germ cell migration. Current Opinion in Cell Biology 42: 128–137. ISSN: 0955-0674. PMID 27484857. PMC 5064876. DOI: 10.1016/j.ceb.2016.07.007.
- ↑ a b c d e f Coffman, Clark R. (May 2003). Cell migration and programmed cell death of Drosophila germ cells. Annals of the New York Academy of Sciences 995 (1): 117–126. ISSN: 0077-8923. PMID 12814944. DOI: 10.1111/j.1749-6632.2003.tb03215.x.
- ↑ Santos, Ana C., Lehmann, Ruth (27 juli 2004). Germ cell specification and migration in Drosophila and beyond. Current Biology 14 (14): R578–589. ISSN: 0960-9822. PMID 15268881. DOI: 10.1016/j.cub.2004.07.018.
- ↑ a b Montell, D. J. (June 1999). The genetics of cell migration in Drosophila melanogaster and Caenorhabditis elegans development. Development 126 (14): 3035–3046. ISSN: 0950-1991. PMID 10375496. DOI: 10.1242/dev.126.14.3035.
- ↑ Gilbert, Scott (2000). Developmental Biology. Sinauer Associates, Sunderland (MA), 100–101. ISBN 0-87893-243-7.
- ↑ (en) Grimaldi, Cecilia, Raz, Erez (April 2020). Germ cell migration—Evolutionary issues and current understanding. Seminars in Cell & Developmental Biology 100: 152–159. PMID 31864795. DOI: 10.1016/j.semcdb.2019.11.015.
- ↑ (en) Richardson, Brian E., Lehmann, Ruth (January 2010). Mechanisms guiding primordial germ cell migration: strategies from different organisms. Nature Reviews Molecular Cell Biology 11 (1): 37–49. ISSN: 1471-0080. PMID 20027186. PMC 4521894. DOI: 10.1038/nrm2815.