Phagocytose

La phagocytose, en biologie, est le processus cellulaire par lequel certaines cellules regroupées sous la dénomination générale de phagocyte peuvent ingérer des particules étrangères solides d'échelle micrométrique. On considère habituellement que la phagocytose est une forme particulière d'endocytose[1]. Elle se distingue d'autres processus d'internalisation cellulaire (comme la pinocytose) par au moins deux critères généraux :
- la phagocytose est induite par le contact physique avec la particule-cible ;
- la particule-cible de la phagocytose a une taille supérieure à 0,5 µm.
Elle est réalisée de façon très efficace par les cellules regroupés sous le terme de cellules phagocytaires professionnelles : iée macrophages, les cellules dendritiques ou les neutrophiles. les ostéoclastes sont aussi des cellules phagocytaires très efficaces. Les fibroblastes, les cellules épithéliales et les cellules endothéliales peuvent également réaliser une phagocytose avec une faible efficacité et sont donc décrits comme des phagocytes non professionnels. Ces cellules ne peuvent pas ingérer de micro-organismes, mais jouent un rôle important dans l’élimination des cellules mortes et le maintien de l’homéostasie [2].
Élément essentiel de l'immunité, elle a été découverte à la fin du XIXe siècle par Élie Metchnikov (1845-1916), récompensé par le Prix Nobel de physiologie ou médecine en 1908, conjointement avec Paul Ehrlich. Chez les amibes, elle joue un rôle nutritionnel, en leur permettant de capturer et d'ingérer des bactéries[1].
Les conséquences de l'activité phagocytaire sont multiples et importantes. On peut citer notamment l'élimination des pathogènes, la présentation des antigènes aux lymphocytes, l'élimination des débris cellulaires pro-inflammatoires. La phagocytose peut également être exploitée en biomédecine, par exemple pour la livraison ciblée de molécules pharmaceutiques ou pour l'immunothérapie antitumorale.
Considérations générales
[modifier | modifier le code]Phagocytes
[modifier | modifier le code]On appelle phagocytes toutes cellules étant capables de phagocytose. Toutefois, la phagocytose est principalement le fait de phagocytes dit « professionnels ». Ce qualificatif leur vient de leur taux de phagocytose nettement plus élevé ainsi que de l'adaptation de leur machinerie moléculaire interne à ce processus d'internalisation. Ces phagocytes « professionnels » appartiennent à la catégorie des leucocytes. On distingue en particulier les macrophages, les neutrophiles et les cellules dendritiques. Les études des bases moléculaires et mécaniques de la phagocytoses concernent principalement les deux premiers.
Diversité morphologique de la phagocytose
[modifier | modifier le code]La phagocytose est habituellement représentée dans la littérature comme la formation d'une protrusion membranaire en forme de calice, la coupe phagocytaire, qui se forme au niveau du contact entre phagocyte et cible phagocytaire. Cependant, les déformations membranaires menant à la phagocytose présentent des morphologies très variées, en fonction de la cible et de la nature des ligands et récepteurs phagocytaires impliqués :
- coupe phagocytaire : c'est la forme la plus classiquement décrite. Elle se forme par exemple en aval de la détection du fragment constant (Fc) des immunoglobulines G (IgG) par les récepteurs au fragment constant (FcR) portes par le phagocyte. Elle constitue en un piédestal, à l'endroit du contact initial entre le phagocyte et la cible, et d'un « calice », la coupe phagocytaire a proprement parler, dont les bords englobent progressivement l’intégralité de la cible ;
- rides membranaires (ruffle) : La phagocytose en aval de la molécule du complément C3bi (détectée par le récepteur au complément CR3) favorise la formation de « rides » membranaires (« ruffles » en anglais) : des ondulations de la membrane plasmique qui peuvent finir par recouvrir la cible phagocytaire ;
- la phagocytose sans protrusions membranaires : des observations microscopiques pionnières ont rapporte des phagocytoses sans protrusions, ou la cible phagocytaire semblait « couler » dans la cellule, par exemple dans le contexte de la phagocytose en aval du complément C3bi, ou dans la phagocytose passive (sans utilisation de récepteurs ou ligands phagocytaires, seulement due a hydrophobie de la cible) ;
- phagocytose « enroulée » : cas particulier, observé lors de la phagocytose de certains pathogènes. La membrane de la cellule s'enroule autour d’elle-même, formant ainsi une spirale ;
Les causes physico-chimiques ainsi que les conséquences fonctionnelles (efficacité, rapidité, etc.) de cette diversité sont encore peu connues.
Diversité des couples récepteurs phagocytaires/ligands phagocytaires
[modifier | modifier le code]La phagocytose se distingue d'autres modes d'ingestion cellulaire par la nécessité d'un contact entre le phagocyte et sa cible. Certains cas de phagocytose passive ont été décrits. Dans ce cas, les propriétés physico-chimique du couple cible/phagocyte (charges électriques, hydrophobie) suffisent à ce que la cible soit passivement englobée. La plupart des cas concernent en revanche une phagocytose active, où le processus n'est enclenché qu'en aval de l'appariement d'un ou plusieurs récepteurs phagocytaires (portés par le phagocyte) et d'un ou plusieurs ligands (portés par la cible). Cette phagocytose active a pour conséquence d'introduire la notion de spécificité de la phagocytose. Parmi les cas de phagocytose active, on distingue ceux où la cible présente déjà les ligands et ceux où les ligands sont fixés sur la cible à la suite de réactions chimiques extérieures, ce qu'on appelle l'opsonisation. L'exemple d'opsonisation le plus connu est le rôle des anticorps IgG : ce n'est que lorsque les IgG se fixent sur leur cible que les récepteurs FcR à la surface des phagocytes peuvent s'y fixer et initier le processus de phagocytose.
Déroulement
[modifier | modifier le code]Détection de la cible
[modifier | modifier le code]La première phase de la phagocytose est la détection de la particule cible. La détection est médiée par des récepteurs dédiés sur les cellules phagocytaires. Les récepteurs de reconnaissance de motifs moléculaires sont les récepteurs de dédiées aux motifs moléculaires associés aux pathogènes. Certains de ces récepteur de reconnaissance de motifs moléculaires peuvent initier la phagocytose. D'autres récepteur de reconnaissance de motifs moléculaires, par exemple les récepteurs de type Toll, peuvent se lier aux motifs moléculaires associés aux pathogènes mais ne pas induire de phagocytose. Ces récepteurs peuvent cependant préparer la cellule à la phagocytose. Enfin Les particules étrangères peuvent également être détectées indirectement par des récepteurs nommés récepteurs opsoniques. Les récepteurs des anticorps et du complément sont les récepteurs opsoniques les mieux décrits. Les autres récepteurs sont appelés récepteurs non opsoniques.
Récepteurs non opsoniques
[modifier | modifier le code]Récepteurs des micro-organismes
[modifier | modifier le code]Les récepteurs qui se lient directement aux motifs moléculaires associés aux pathogènes et peuvent induire une phagocytose comprennent la dectine-1, Mincle (Macrophage inducible Ca2+-dependent lectin receptor), MCL (Macrophage C-type lectin) et DC-SIGN ((Dendritic Cell-Specific Intercellular adhesion molecule-3-Grabbing Non-integrin) ) . Toutes ces molécules font partie de la famille des récepteurs aux lectines de type C [3].
La dectine-1 reconnaît les polysaccharides de levure et s'est avérée être un véritable récepteur phagocytaire. Lorsqu'elle est exprimée sur des cellules hétérologues non phagocytaires, la dectine-1 permet aux cellules d'effectuer la phagocytose [4]. La dectine-1 coopère aussi avec d'autres récepteurs phagocytaires: chez les neutrophiles, elle se connecte au récepteur phagocytaire Mac-1 [5].
Le Mincle (lectine de type C inductible par les macrophages) est un récepteur du dimycolate de tréhalose (TDM), présent sur la paroi cellulaire de certaines mycobactéries [6]. MCL (lectine de type C du macrophage, Dectin-3) est un autre récepteur du TDM qui se lie également aux α-mannanes. Mincle et MCL sont tous deux considérés comme de véritables récepteurs phagocytaires . Dans les cellules myéloïdes, Mincle et MCL semblent coopérer pour améliorer la phagocytose en formant des hétérodimères sur la membrane cellulaire [5].
DC-SIGN est un autre récepteur qui peut se lier à plusieurs agents pathogènes microbiens, notamment des virus, des champignons et des bactéries [3], grâce à la reconnaissance des glycanes fucosylés et des glycanes riches en mannose [7]. DC-SIGN s'est révélé être un récepteur phagocytaire en l'exprimant dans des cellules myéloïdes humaines K562 non phagocytaires ou dans des cellules épithéliales HeLa. DC-SIGNR est un autre récepteur de lectine de type C présentant une forte homologie avec DC-SIGN et capable de se lier à des ligands riches en mannose [8].
Récepteurs des cellules apoptiques
[modifier | modifier le code]Dans les organismes multicellulaires, de nombreuses cellules meurent constamment par apoptose pour maintenir l'homéostasie. Ces cellules apoptotiques sont éliminées par phagocytose. La détection des cellules apoptotiques nécessite des récepteurs particuliers pour des molécules qui n'apparaissent que sur la membrane des cellules mourantes. Ces molécules comprennent la lysophosphatidylcholine et la phosphatidylsérine [9]. Ces molécules délivrent aux phagocytes un signal « mange-moi » [10]. Les récepteurs reconnaissant directement la phosphatidylsérine comprennent la TIM-1 (« T-cell immunoglobulin and mucin domain 1 ») ,la TIM-4 (« T-cell immunoglobulin and mucin domain 4 ») [11], la stabiline-2 [12] et BAI-1 (« Brain-specific angiogenesis inhibitor 1 ») [13].
Certaines cellules normales, par exemple les lymphocytes B et T activés, peuvent exprimer des taux importants de phosphatidylsérine à leur surface. Ces cellules évitent la phagocytose en exprimant en même temps des molécules qui servent de signaux « ne me mange pas » [2]. L’une de ces molécules est le CD47, un ligand du récepteur SIRPα (« Signal regulatory protein α »), qui est exprimé sur les phagocytes [14].
Récepteurs opsoniques
[modifier | modifier le code]Récepteurs d'immunoglobuline
[modifier | modifier le code]
Les récepteurs Fcγ (FcγR) sont des glycoprotéines qui se lient spécifiquement à la chaine lourde (partie Fc) des molécules d'immunoglobulines G [15]. Lorsque les FcγR engagent des molécules d'immunoglobulines G dans des complexes multivalents antigène-anticorps, elles se regroupent sur la membrane de la cellule, puis déclenchent la phagocytose ainsi que d'autres réponses cellulaires. Trois types de FcγR sont exprimés sur les cellules humaines, FcγRI (CD64), FcγRII (CD32) et FcγRIII (CD16) [16].
FcγRI possède trois domaines de type Ig et présente une haute affinité pour les molécules d'immunoglobulines G. En revanche, FcγRII et FcγRIII possèdent deux domaines de type Ig et affichent une faible affinité pour les molécules d'immunoglobulines G [17].
FcγRII présente deux isoformes : FcγRIIa exprimé principalement dans les cellules phagocytaires et FcγRIIb exprimé principalement dans les lymphocytes B [16]. FcγRIIb fonctionne comme un régulateur négatif de la phagocytose [18].
FcγRIII présente deux isoformes : FcγRIIIa exprimé dans les macrophages, les cellules tueuses naturelles , les basophiles, les mastocytes et les cellules dendritiques, et FcγRIIIb exprimé exclusivement sur les neutrophiles [17]. FcγRIIIa est un récepteur avec une partie transmembranaire et une queue cytoplasmique, associé à un dimère de chaînes FcRγ, tandis que FcγRIIIb est un récepteur lié au glycosylphosphatidylinositol , dépourvu de queue cytoplasmique et d'aucune sous-unité associée connue [19].
Récepteurs du complément
[modifier | modifier le code]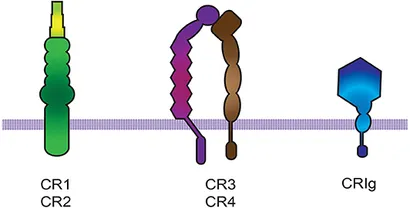
Les molécules du complément déposées sur les micro-organismes ou les cellules se lient au récepteurs du complément de la cellule phagocytaires [20]. Les récepteurs du complément appartiennent à trois groupes de molécules : CR1 (Complement receptor 1) et CR2 (Complement receptor 2), CR3 et CR4, qui appartiennent à la famille des intégrines β2 [21], et CRIg (Complement receptor of the immunoglobulin family), qui appartient à la superfamille des immunoglobulines [22]. L'intégrine αMβ2 (également connue sous le nom de CD11b/CD18, CR3 ou Mac-1) constitue le récepteur phagocytaire le plus efficace parmi les récepteurs du complément [21].
Coordination entre récepteurs
[modifier | modifier le code]Pour une reconnaissance efficace de la cible, de nombreux récepteurs phagocytaires sur la membrane doivent interagir avec plusieurs molécules d’immunoglobulines sur la particule opsonisée. Pour cela, les récepteurs doivent avoir une bonne mobilité membranaire pour pouvoir s'agréger et s'activer [23]. La mobilité n’est pas facile pour la plupart des récepteurs phagocytaires, car ils font partie d’autres glycoprotéines transmembranaires (généralement plus grosses) qui recouvrent la surface cellulaire. Les récepteurs phagocytaires sont des petites molécules qui sont masqués par une couche de grosses glycoprotéines (le glycocalyx), telles que les mucines, l'hyaluronane et les phosphatases membranaires CD45 et CD148 [24]. Enfin, de nombreuses grosses glycoprotéines sont liées au cytosquelette et peuvent interférer avec la diffusion latérale des récepteurs sur la membrane cellulaire [25].
Activation de la formation du phagosome
[modifier | modifier le code]Lorsqu'une particule est reconnue par les récepteurs phagocytaires, diverses voies de signalisation sont activées pour initier la phagocytose. Une réorganisation du cytosquelette d'actine et des modifications de la membrane ont lieu, entraînant une dépression de la zone membranaire touchant la particule et formant la coupe phagocytaire. Ensuite, des pseudopodes se forment autour de la particule jusqu'à ce que la membrane recouvre complètement la particule pour former un nouveau phagosome à l'intérieur de la cellule. Les mécanismes de signalisation permettant d'activer la phagocytose sont mieux connus pour les récepteurs Fc et les récepteurs du complément . Pour d’autres récepteurs phagocytaires, les voies de signalisation commencent tout juste à être étudiées. Dépendant du type cellulaire et des conditions de phagocytose, des sources autres que la membrane plasmique peuvent fournir la membrane du phagosome. Il s'agit des endosomes de recyclage et du réticulum endoplasmique.
Voie d'activation des récepteurs Fc
[modifier | modifier le code]Voie d'activation des récepteurs du complément
[modifier | modifier le code]
Parmi les récepteurs du complément, le CR3 est le récepteur phagocytaire le plus efficace [21]. Dès les premières études, il a été réalisé que le CR3 sur les macrophages initie un type de phagocytose différent de celui médié par les récepteurs des anticorps Fcγ. La phagocytose médiée par CR3 est caractérisée par « l'enfoncement » de la particule cible dans la membrane cellulaire sans génération de pseudopodes autour de la particule [31]. En outre, l’utilisation des composants du cytosquelette pour l’internalisation des particules est différente entre la phagocytose médiée par les récepteurs aux immunoglobulines et les récepteurs du complément. Au cours de la phagocytose médiée par les récepteurs aux immunoglobulines, le cytosquelette d'actine est utilisé seul , tandis que lors de la phagocytose médiée par les récepteurs du complément, les cytosquelettes d'actine et de microtubules sont impliqués [29],[30]. Dans la phagocytose du complément, le remodelage de l'actine F dépend de l'activation de la GTPase Rho, mais pas des GTPases Rac ou Cdc42 comme pour l'activation par les récepteurs des immunoglobulines [32]. La GTPase Rho, à son tour, favorise la polymérisation de l'actine via deux mécanismes (Voir vignette).
Formation du phagosome
[modifier | modifier le code]
La phagocytose démarre lorsque les récepteurs phagocytaires engagent des ligands sur la particule à ingérer. Ensuite, les récepteurs activent des voies de signalisation qui modifient la composition de la membrane et contrôlent le cytosquelette d'actine, entraînant la formation de protubérances membranaires destinées à recouvrir la particule. Enfin, ces saillies membranaires fusionnent au niveau distal, créant une nouvelle vésicule qui se détache de la membrane plasmique. Cette nouvelle vésicule contenant la particule ingérée est le phagosome [38].
Lors de la formation des phagosomes, la membrane change sa composition lipidique impliquant la formation et la dégradation de différentes molécules lipidiques sur la membrane du phagosome de manière ordonnée [39].
Au cours de la phagocytose médiée par le récepteur Fcγ, le phosphatidylinositol-4,5-bisphosphate s'accumule initialement au niveau de la saillie phagocytaire, mais diminue ensuite rapidement [40]. La diminution de phosphatidylinositol-4,5-bisphosphate est importante pour l'internalisation des particules, probablement en facilitant le désassemblage de l'actine . Cette diminution est provoquée par l'action de la phosphoinositide 3-kinase, qui le phosphoryle pour produire du Phosphatidylinositol-3,4,5-trisphosphate 3-phosphatase au niveau de la saillie phagocytaire [41]. La réduction du phosphatidylinositol-4,5-bisphosphate dans la membrane est également médiée par l'action de la phospholipase Cγ [42].
Parallèlement aux changements dans la composition lipidique, la membrane plasmique change également en remodelant le cytosquelette d'actine afin de générer les saillies membranaires qui recouvriront la particule cible. Premièrement, le cytosquelette cortical est perturbé. Deuxièmement, les pseudopodes sont formés par polymérisation de la F-actine. Troisièmement, à la base de la saillie phagocytaire, l'actine est dépolymérisée tandis que le phagosome membranaire est scellé à l'extrémité distale pour former le phagosome [43].
Phagosome
[modifier | modifier le code]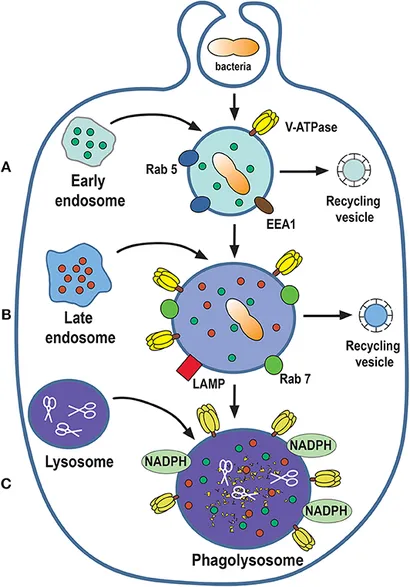
Une fois dans la cellule phagocytaire, , le nouveau phagosome transforme sa composition membranaire et son contenu, pour devenir une nouvelle vésicule, le phagolysosome, capable de dégrader la particule ingérée. Cette transformation est connue sous le nom de maturation du phagosome et consiste en des interactions successives de fusion et de fission entre le nouveau phagosome et les endosomes précoces, les endosomes tardifs et enfin les lysosomes [44].
Dépendant du type cellulaire et des conditions de phagocytose, des sources autres que la membrane plasmique peuvent fournir la membrane du phagosome. Il s'agit des endosomes de recyclage et du réticulum endoplasmique[45].
Digestion
[modifier | modifier le code]À l'intérieur de ce phagosome, le devenir de la particule peut être de trois types :
- la digestion consécutive à l'accolement et à la fusion des lysosomes avec la membrane du phagosome constituant ainsi un phagolysosome duquel les divers enzymes vont se déverser et, selon leur spécificité, s'attaquer aux divers constituants de la particule ou du micro-organisme ;
- la persistance est observée avec les particules difficilement biodégradables, mais également avec certains germes ;
- la multiplication intracytoplasmique (relatif à l'intérieur du cytoplasme), qui est le propre de certaines variétés de parasites ou de virus, de bactéries responsables d'infections particulièrement virulentes comme la brucellose et la tuberculose.
La phagocytose s’accompagne le plus souvent d’une brutale élévation de la consommation cellulaire en oxygène. En effet, la liaison d’une particule exogène à son récepteur membranaire entraîne une activation de la NADPH oxydase, qui transfère des électrons du NADPH à l'oxygène moléculaire. Les formes actives de l'oxygène formées (Peroxyde d'hydrogène H2O2, radicaux OH• et O2) se retrouvent en forte concentration dans le phagosome, où elles exercent des effets toxiques sur les micro-organismes phagocytés. De plus, certains macrophages sont capables après induction d’une NO synthase de produire des dérivés nitrés, notamment le monoxyde d'azote, par oxydation des noyaux azotés de la L-arginine. Le monoxyde d’azote, qui est libéré lors de l'infection de macrophages par des bactéries, des levures, des parasites ou des champignons, contribue à l'élimination de ces organismes.
Pendant l’étape de digestion, il y a l’activation de lysosomes, constituant ainsi un phagolysosome. Les divers enzymes vont se déverser et, selon leur spécificité, s'attaquer aux divers constituants de la particule ou du micro-organisme pour la digestion totale.
Mécanisme d' échappement microbien à la phagocytose et conséquences biologiques
[modifier | modifier le code]La phagocytose est un processus efficace qui se termine par la formation du phagolysosome et son environnement très hostile pour la plupart des micro-organismes. Il n’est donc pas surprenant que de nombreux agents pathogènes efficaces aient développé de multiples stratégies pour prévenir et/ou empêcher la phagocytose. Ces stratégies incluent la prévention de la phagocytose, l'interférence de la maturation du phagosome, la résistance au contenu du phagolysosome et même la fuite physique du phagosome. Ces phénomènes d'échappement ont particulièrement étudié chez le staphylocoque dorée, M. tuberculosis et L. monocytogenes.
Dans les cas de persistances par des micro-organismes (levures, bactéries ou virus), il existe la digestion partielle avec présentation des peptides antigéniques par les molécules de classe II du CMH.
La multiplication du micro-organisme :
- qui empêche la fusion des lysosomes (Listeria, Legionella, Chlamydia, Mycobacterium tuberculosis, Toxoplasma) ;
- qui survit dans le phagolysosome (Salmonella, Histoplasma) ;
- qui sort du phagosome vers le cytoplasme où il se multiplie (Listeria, Shigella).
Dans tous les cas, le macrophage doit être activé : c'est le rôle de la cellule Th1 qui, par la sécrétion d'interférons, activera le macrophage « embarrassé ».
Prévention de la phagocytose
[modifier | modifier le code]Perturbation dans la formation du phagosome
[modifier | modifier le code]Résistance au contenu du phagolysosome
[modifier | modifier le code]Échappement physique du phagolysosome
[modifier | modifier le code]Méthodes classiques d'étude de la phagocytose
[modifier | modifier le code]Notes et références
[modifier | modifier le code]- Cet article est une traduction partielle de Uribe-Querol E and Rosales C (2020) Phagocytosis: Our Current Understanding of a Universal Biological Process. Front. Immunol. 11:1066. doi: 10.3389/fimmu.2020.01066
- Royet, « Endocytose 2-2 1.ppt », sur biologie.univ-mrs.fr, (consulté le ).
- (en) Siamon Gordon, « Phagocytosis: An Immunobiologic Process », Immunity, vol. 44, no 3, , p. 463–475 (DOI 10.1016/j.immuni.2016.02.026, lire en ligne, consulté le )
- Li K, Underhill DM. C-type lectin receptors in phagocytosis. In: Current Topics in Microbiology and Immunology. Berlin; Heidelberg: Springer Nature. (2020). p. 18
- (en) J. Herre, « Dectin-1 uses novel mechanisms for yeast phagocytosis in macrophages », Blood, vol. 104, no 13, , p. 4038–4045 (ISSN 0006-4971 et 1528-0020, DOI 10.1182/blood-2004-03-1140, lire en ligne, consulté le )
- (en) Xun Li, Ahmad Utomo, Xavier Cullere et Myunghwan Mark Choi, « The β-Glucan Receptor Dectin-1 Activates the Integrin Mac-1 in Neutrophils via Vav Protein Signaling to Promote Candida albicans Clearance », Cell Host & Microbe, vol. 10, no 6, , p. 603–615 (PMID 22177564, PMCID PMC3244687, DOI 10.1016/j.chom.2011.10.009, lire en ligne, consulté le )
- (en) Eri Ishikawa, Tetsuaki Ishikawa, Yasu S. Morita et Kenji Toyonaga, « Direct recognition of the mycobacterial glycolipid, trehalose dimycolate, by C-type lectin Mincle », Journal of Experimental Medicine, vol. 206, no 13, , p. 2879–2888 (ISSN 1540-9538 et 0022-1007, PMID 20008526, PMCID PMC2806462, DOI 10.1084/jem.20091750, lire en ligne, consulté le )
- (en) Ellis van Liempt, Christine M.C. Bank, Padmaja Mehta et Juan Jesús Garcı´a-Vallejo, « Specificity of DC‐SIGN for mannose‐ and fucose‐containing glycans », FEBS Letters, vol. 580, no 26, , p. 6123–6131 (ISSN 0014-5793 et 1873-3468, DOI 10.1016/j.febslet.2006.10.009, lire en ligne, consulté le )
- (en) Chae Gyu Park, Kazuhiko Takahara, Eiji Umemoto et Yusuke Yashima, « Five mouse homologues of the human dendritic cell C-type lectin, DC-SIGN », International Immunology, vol. 13, no 10, , p. 1283–1290 (ISSN 1460-2377 et 0953-8178, DOI 10.1093/intimm/13.10.1283, lire en ligne, consulté le )
- (en) S Nagata, J Suzuki, K Segawa et T Fujii, « Exposure of phosphatidylserine on the cell surface », Cell Death & Differentiation, vol. 23, no 6, , p. 952–961 (ISSN 1350-9047 et 1476-5403, PMID 26891692, PMCID PMC4987739, DOI 10.1038/cdd.2016.7, lire en ligne, consulté le )
- (en) Katsumori Segawa et Shigekazu Nagata, « An Apoptotic ‘Eat Me’ Signal: Phosphatidylserine Exposure », Trends in Cell Biology, vol. 25, no 11, , p. 639–650 (DOI 10.1016/j.tcb.2015.08.003, lire en ligne, consulté le )
- (en) Norimoto Kobayashi, Piia Karisola, Victor Peña-Cruz et David M. Dorfman, « TIM-1 and TIM-4 Glycoproteins Bind Phosphatidylserine and Mediate Uptake of Apoptotic Cells », Immunity, vol. 27, no 6, , p. 927–940 (PMID 18082433, PMCID PMC2757006, DOI 10.1016/j.immuni.2007.11.011, lire en ligne, consulté le )
- (en) S-Y Park, M-Y Jung, H-J Kim et S-J Lee, « Rapid cell corpse clearance by stabilin-2, a membrane phosphatidylserine receptor », Cell Death & Differentiation, vol. 15, no 1, , p. 192–201 (ISSN 1350-9047 et 1476-5403, DOI 10.1038/sj.cdd.4402242, lire en ligne, consulté le )
- (en) Daeho Park, Annie-Carole Tosello-Trampont, Michael R. Elliott et Mingjian Lu, « BAI1 is an engulfment receptor for apoptotic cells upstream of the ELMO/Dock180/Rac module », Nature, vol. 450, no 7168, , p. 430–434 (ISSN 0028-0836 et 1476-4687, DOI 10.1038/nature06329, lire en ligne, consulté le )
- (en) Richard K. Tsai et Dennis E. Discher, « Inhibition of “self” engulfment through deactivation of myosin-II at the phagocytic synapse between human cells », The Journal of Cell Biology, vol. 180, no 5, , p. 989–1003 (ISSN 1540-8140 et 0021-9525, PMID 18332220, PMCID PMC2265407, DOI 10.1083/jcb.200708043, lire en ligne, consulté le )
- Carlos Rosales et Eileen Uribe-Querol, « Fc receptors: Cell activators of antibody functions », Advances in Bioscience and Biotechnology, vol. 04, no 04, , p. 21–33 (ISSN 2156-8456 et 2156-8502, DOI 10.4236/abb.2013.44A004, lire en ligne, consulté le )
- (en) Jeffrey V. Ravetch et Silvia Bolland, « IgG Fc Receptors », Annual Review of Immunology, vol. 19, no 1, , p. 275–290 (ISSN 0732-0582 et 1545-3278, DOI 10.1146/annurev.immunol.19.1.275, lire en ligne, consulté le )
- Ravetch JV. Fc receptors. In: Paul WE, editor. Fundamental Immunology. Philadelphia: Lippincott Williams & Wilkins (2003). p. 631–84.
- (en) Susheela Tridandapani, Kristina Siefker, Jo Ellen Carter et Mark D. Wewers, « Regulated Expression and Inhibitory Function of FcγRIIb in Human Monocytic Cells », Journal of Biological Chemistry, vol. 277, no 7, , p. 5082–5089 (DOI 10.1074/jbc.M110277200, lire en ligne, consulté le )
- (en) Lisa C. Willcocks, Kenneth G.C. Smith et Menna R. Clatworthy, « Low-affinity Fcγ receptors, autoimmunity and infection », Expert Reviews in Molecular Medicine, vol. 11, (ISSN 1462-3994, DOI 10.1017/S1462399409001161, lire en ligne, consulté le )
- (en) Eric Brown, « Complement Receptors, Adhesion, and Phagocytosis », dans Molecular Mechanisms of Phagocytosis, Springer US, , 49–57 p. (ISBN 978-0-387-25419-7, DOI 10.1007/978-0-387-28669-3_4, lire en ligne)
- (en) Michael L. Dustin, « Complement Receptors in Myeloid Cell Adhesion and Phagocytosis », Microbiology Spectrum, vol. 4, no 6, (ISSN 2165-0497, PMID 27809953, PMCID PMC5580235, DOI 10.1128/microbiolspec.MCHD-0034-2016, lire en ligne, consulté le )
- (en) Menno van Lookeren Campagne, Christian Wiesmann et Eric J. Brown, « Macrophage complement receptors and pathogen clearance », Cellular Microbiology, vol. 9, no 9, , p. 2095–2102 (DOI 10.1111/j.1462-5822.2007.00981.x, lire en ligne, consulté le )
- (en) Valentin Jaumouillé et Sergio Grinstein, « Receptor mobility, the cytoskeleton, and particle binding during phagocytosis », Current Opinion in Cell Biology, vol. 23, no 1, , p. 22–29 (DOI 10.1016/j.ceb.2010.10.006, lire en ligne, consulté le )
- (en) Timothy A. Springer, « Adhesion receptors of the immune system », Nature, vol. 346, no 6283, , p. 425–434 (ISSN 0028-0836 et 1476-4687, DOI 10.1038/346425a0, lire en ligne, consulté le )
- (en) Spencer A. Freeman et Sergio Grinstein, « Phagocytosis: receptors, signal integration, and the cytoskeleton », Immunological Reviews, vol. 262, no 1, , p. 193–215 (ISSN 0105-2896 et 1600-065X, DOI 10.1111/imr.12212, lire en ligne, consulté le )
- (en) Isabel M. Olazabal, Emmanuelle Caron, Robin C. May et Kerstin Schilling, « Rho-Kinase and Myosin-II Control Phagocytic Cup Formation during CR, but Not FcγR, Phagocytosis », Current Biology, vol. 12, no 16, , p. 1413–1418 (DOI 10.1016/S0960-9822(02)01069-2, lire en ligne, consulté le )
- (en) Emma Colucci-Guyon, Florence Niedergang, Bradley J. Wallar et Jun Peng, « A Role for Mammalian Diaphanous-Related Formins in Complement Receptor (CR3)-Mediated Phagocytosis in Macrophages », Current Biology, vol. 15, no 22, , p. 2007–2012 (DOI 10.1016/j.cub.2005.09.051, lire en ligne, consulté le )
- (en) Elodie Lewkowicz, Floriane Herit, Christophe Le Clainche et Pierre Bourdoncle, « The microtubule-binding protein CLIP-170 coordinates mDia1 and actin reorganization during CR3-mediated phagocytosis », The Journal of Cell Biology, vol. 183, no 7, , p. 1287–1298 (ISSN 1540-8140 et 0021-9525, PMID 19114595, PMCID PMC2606960, DOI 10.1083/jcb.200807023, lire en ligne, consulté le )
- (en) L A Allen et A Aderem, « Molecular definition of distinct cytoskeletal structures involved in complement- and Fc receptor-mediated phagocytosis in macrophages. », The Journal of experimental medicine, vol. 184, no 2, , p. 627–637 (ISSN 0022-1007 et 1540-9538, PMID 8760816, PMCID PMC2192718, DOI 10.1084/jem.184.2.627, lire en ligne, consulté le )
- Newman SL, Mikus LK, Tucci MA. Differential requirements for cellular cytoskeleton in human macrophage complement receptor- and Fc receptor-mediated phagocytosis. J Immunol. (1991) 146:967–74.
- (en) Alan Aderem et David M. Underhill, « MECHANISMS OF PHAGOCYTOSIS IN MACROPHAGES », Annual Review of Immunology, vol. 17, no 1, , p. 593–623 (ISSN 0732-0582 et 1545-3278, DOI 10.1146/annurev.immunol.17.1.593, lire en ligne, consulté le )
- (en) Emmanuelle Caron et Alan Hall, « Identification of Two Distinct Mechanisms of Phagocytosis Controlled by Different Rho GTPases », Science, vol. 282, no 5394, , p. 1717–1721 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.282.5394.1717, lire en ligne, consulté le )
- (en) Carlos Rosales et Eileen Uribe-Querol, « Phagocytosis: A Fundamental Process in Immunity », BioMed Research International, vol. 2017, , p. 1–18 (ISSN 2314-6133 et 2314-6141, PMID 28691037, PMCID PMC5485277, DOI 10.1155/2017/9042851, lire en ligne, consulté le )
- (en) Adam D. Hoppe et Joel A. Swanson, « Cdc42, Rac1, and Rac2 Display Distinct Patterns of Activation during Phagocytosis », Molecular Biology of the Cell, vol. 15, no 8, , p. 3509–3519 (ISSN 1059-1524 et 1939-4586, PMID 15169870, PMCID PMC491814, DOI 10.1091/mbc.e03-11-0847, lire en ligne, consulté le )
- (en) Yingyu Mao et Silvia C Finnemann, « Regulation of phagocytosis by Rho GTPases », Small GTPases, vol. 6, no 2, , p. 89–99 (ISSN 2154-1248 et 2154-1256, PMID 25941749, PMCID PMC4601285, DOI 10.4161/21541248.2014.989785, lire en ligne, consulté le )
- (en) Nobukazu Araki, Tanenori Hatae, Aizo Furukawa et Joel A. Swanson, « Phosphoinositide-3-kinase-independent contractile activities associated with Fcγ-receptor-mediated phagocytosis and macropinocytosis in macrophages », Journal of Cell Science, vol. 116, no 2, , p. 247–257 (ISSN 1477-9137 et 0021-9533, DOI 10.1242/jcs.00235, lire en ligne, consulté le )
- (en) Anna E. Dart, Sylvain Tollis, Michael D. Bright et Gad Frankel, « The motor protein myosin 1G functions in FcγR-mediated phagocytosis », Journal of Cell Science, vol. 125, no 24, , p. 6020–6029 (ISSN 1477-9137 et 0021-9533, DOI 10.1242/jcs.109561, lire en ligne, consulté le )
- Eileen Uribe-Querol et Carlos Rosales, « Phagocytosis: Our Current Understanding of a Universal Biological Process », Frontiers in Immunology, vol. 11, (ISSN 1664-3224, PMID 32582172, PMCID PMC7280488, DOI 10.3389/fimmu.2020.01066, lire en ligne, consulté le )
- (en) Roni Levin, Sergio Grinstein et Johnathan Canton, « The life cycle of phagosomes: formation, maturation, and resolution », Immunological Reviews, vol. 273, no 1, , p. 156–179 (ISSN 0105-2896 et 1600-065X, DOI 10.1111/imr.12439, lire en ligne, consulté le )
- (en) Roberto J. Botelho, Mary Teruel, Renee Dierckman et Richard Anderson, « Localized Biphasic Changes in Phosphatidylinositol-4,5-Bisphosphate at Sites of Phagocytosis », The Journal of Cell Biology, vol. 151, no 7, , p. 1353–1368 (ISSN 0021-9525 et 1540-8140, PMID 11134066, PMCID PMC2150667, DOI 10.1083/jcb.151.7.1353, lire en ligne, consulté le )
- (en) John G. Marshall, James W. Booth, Vuk Stambolic et Tak Mak, « Restricted Accumulation of Phosphatidylinositol 3-Kinase Products in a Plasmalemmal Subdomain during Fcγ Receptor-Mediated Phagocytosis », The Journal of Cell Biology, vol. 153, no 7, , p. 1369–1380 (ISSN 0021-9525 et 1540-8140, PMID 11425868, PMCID PMC2150726, DOI 10.1083/jcb.153.7.1369, lire en ligne, consulté le )
- (en) Elaine C. Larsen, Jeannine A. DiGennaro, Naoaki Saito et Sapna Mehta, « Differential Requirement for Classic and Novel PKC Isoforms in Respiratory Burst and Phagocytosis in RAW 264.7 Cells », The Journal of Immunology, vol. 165, no 5, , p. 2809–2817 (ISSN 0022-1767 et 1550-6606, DOI 10.4049/jimmunol.165.5.2809, lire en ligne, consulté le )
- (en) Spencer A. Freeman et Sergio Grinstein, « Phagocytosis: receptors, signal integration, and the cytoskeleton », Immunological Reviews, vol. 262, no 1, , p. 193–215 (ISSN 0105-2896 et 1600-065X, DOI 10.1111/imr.12212, lire en ligne, consulté le )
- (en) Gregory D. Fairn et Sergio Grinstein, « How nascent phagosomes mature to become phagolysosomes », Trends in Immunology, vol. 33, no 8, , p. 397–405 (DOI 10.1016/j.it.2012.03.003, lire en ligne, consulté le )
- Nature Reviews Immunology 3, 280-291 (2003)
Voir aussi
[modifier | modifier le code]Articles connexes
[modifier | modifier le code]Liens externes
[modifier | modifier le code]- Notices dans des dictionnaires ou encyclopédies généralistes :
- Ressources relatives à la santé :