Cerveau humain

| Système | |
|---|---|
| Vascularisation | |
| Drainage veineux | |
| Embryologie | |
| Comprend |
| Nom latin |
Cerebrum |
|---|---|
| TA98 |
A14.1.03.001 |
| TA2 |
5415 |
| FMA |
50801 |
| Sujet |
184 |
|---|---|
| Page |
736 |
Le cerveau humain a la même structure générale que le cerveau des autres mammifères, mais il est celui dont la taille relative par rapport au reste du corps est devenue la plus grande au cours de l'évolution. Le cerveau humain est la structure vivante la plus complexe que nous connaissons. Cet organe n'est pas homogène et sa complexité est représentée par la juxtaposition de différents territoires dont les fonctions sont spécifiées[1]
Si la baleine bleue a le cerveau le plus lourd avec 6,92 kilogrammes contre environ 1,5 kg pour celui de l'homme, le coefficient d'encéphalisation humain est le plus élevé et est sept fois supérieur à celui de la moyenne des mammifères[2]. L'augmentation du volume cérébral humain vient en grande partie du développement du cortex cérébral qui est bien distinct de celui des autres Primates, en particulier les lobes frontaux qui représentent plus de 30 % de la surface cérébrale et interviennent essentiellement dans la planification, le langage et le mouvement volontaire. Près de la moitié du cortex cérébral est consacré à l'analyse sensorielle, principalement la vision[3].
Bien qu'il soit protégé par la barrière hémato-encéphalique et les os épais du crâne et qu'il baigne dans le liquide cérébrospinal, le cerveau humain reste soumis à des blessures et maladies dont les plus fréquentes sont le traumatisme crânien, les maladies neurotoxiques, neurologiques et neurodégénératives. Un certain nombre de troubles psychiques, comme la schizophrénie et la dépression, sont considérés comme associés à des dysfonctionnements du cerveau, bien que la cause de ces troubles soit mal comprise.
Histoire de la connaissance du cerveau humain
[modifier | modifier le code]Historiquement, les avis se sont souvent opposés pour savoir qui du cerveau ou du cœur était le siège de l'âme. Dans un sens, il était impossible de nier que la conscience semble comme localisée dans la tête, qu'un coup à la tête provoque bien plus facilement l'inconscience qu'un coup sur le torse, et que secouer la tête donne des vertiges. Dans un autre sens, le cerveau soumis à un examen superficiel semble inerte, tandis que le cœur bat constamment. L'arrêt des battements cardiaques provoque la mort, tandis que les émotions induisent des changements du rythme cardiaque et que le chagrin produit souvent une sensation de douleur au niveau du cœur (« cœur brisé »). Pour Aristote, le siège de l'âme était le cœur. Le cerveau, organe de refroidissement, servait simplement à assurer la circulation du sang (théorie philosophique et médicale du cardiocentrisme). Démocrite divise l'âme en trois parties : l'intellect dans la tête, l'émotion dans le cœur, le désir autour du foie[4]. Hippocrate était sûr que l'âme se trouvait dans le cerveau (théorie philosophique et médicale du cérébrocentrisme ou céphalocentrisme). Pour Hérophile, le cerveau était le centre de l’intelligence[5].

Galien a aussi défendu l'importance du cerveau, et élaboré des théories assez avancées sur sa manière de fonctionner. Même après la reconnaissance de la primauté du cerveau par la société savante, l'idée du cœur comme siège de l'intelligence a continué de survivre dans les idiomes populaires[6]. Galien a entrepris un long travail pour décrire les relations anatomiques entre le cerveau, les nerfs, et les muscles, démontrant que tous les muscles du corps sont connectés au cerveau par un réseau de nerfs. Il postulait que les nerfs activent les muscles mécaniquement, en transportant une substance mystérieuse qu'il appelle le pneuma psychique (ou « esprit animal »). Ses idées connaissent une relative renommée pendant le Moyen Âge, mais ne progressent pas jusqu'à la Renaissance. À la Renaissance, des études anatomiques détaillées résument et combinent les idées de Galien avec celles de Descartes et ses successeurs. Comme Galien, Descartes pensait le système nerveux en termes hydrauliques. Il est convaincu que les plus hautes fonctions cognitives, le langage notamment, sont effectuées par une « chose pensante » non-physique (res cogitans), mais que la majorité des comportements humains et animaux peuvent s'expliquer mécaniquement. Le premier grand pas vers la médecine moderne provient des recherches de Luigi Galvani, qui découvre qu'une décharge d'électricité statique appliquée sur le nerf d'une grenouille morte provoque la contraction de sa jambe.
Chaque grande avancée dans la compréhension du cerveau a suivi plus ou moins directement le développement d'une nouvelle méthode d'analyse.
Au XVIIe siècle, Thomas Willis, père des neurosciences modernes, prend le contrepied de la théorie dualiste de Descartes, privilégiant l'approche matérialiste grâce à sa description du fonctionnement des nerfs. Préoccupé par la recherche du lien entre cerveau et esprit, Willis étend les concepts proposés par Galien, selon lesquels le cerveau était l'organe responsable de l'excrétion des « esprits animaux » (théorie ventriculaire des cerveaux), et situe la pensée dans le cortex cérébral.
Au XVIIIe siècle, Vincenzo Malacarne (it), fondateur de l'anatomie topographique et de la neuropsychologie, pose les principes de l'existence de signes cliniques de ses patients reliés à l’observation post-mortem de lésions de leur cerveau. Paul Broca démontre cette corrélation anatomoclinique[7].
Au XIXe siècle, Franz Gall développe grâce à des techniques organologiques, une théorie localisationiste selon laquelle les fonctions cognitives sont au niveau des circonvolutions du cortex cérébral[8]. Les chirurgiens Charles Scott Sherrington et C.S. Roy observent le cerveau en salle d'opération et remarquent qu'une zone du cortex rougit lorsque le patient bouge une partie précise de son corps, montrant la consommation d'oxygène cérébrale.
Jusqu'au début du XXe siècle, les principales avancées ont découlé de la découverte de nouveaux colorants histologiques[9]. Sans coloration, le tissu cérébral apparait au microscope comme un impénétrable enchevêtrement de fibres protoplasmiques dans lequel il était impossible de déterminer une quelconque structure. L'une des colorations les plus importantes est la coloration de Golgi qui permet de colorer seulement une petite fraction de neurones. La coloration des neurones se fait apparemment de façon aléatoire mais colore les neurones dans leur entier, aussi bien le corps cellulaire que les dendrites et l'axone. Grâce notamment aux travaux de Camillo Golgi et de Santiago Ramón y Cajal, la nouvelle coloration a pu révéler des centaines de types distincts de neurones, chacun avec sa propre structure dendritique et son propre schéma de connectivité.
La mesure du rapport des propriétés des organismes selon leur taille et leur masse est initiée dans les années 1920 par le biologiste Julian Huxley, qui la nomme allométrie.
Au XXe siècle, les progrès en électronique ont ouvert la voie à la recherche des propriétés électriques des cellules nerveuses, notamment aux travaux comme ceux de Louis Sokoloff sur l'autoradiographie (le débit sanguin cérébral par la méthode des traceurs radioactifs permet de réaliser une cartographie fonctionnelle[10]), d'Alan Hodgkin ou d'Andrew Huxley sur la biophysique du potentiel d'action, et les travaux sur l'électrochimie de la synapse comme ceux de Bernard Katz[11]. Les premières études utilisaient des préparations spéciales, comme avec l'étude de la réponse de fuite rapide du calmar qui fait intervenir un axone et des synapses géantes. L'amélioration constante des électrodes et de l'électronique a permis d'affiner le niveau de résolution des recherches. Ces études ont complété les planches anatomiques en concevant le cerveau comme une entité dynamique.
Aux États-Unis, les années 1990 sont déclarées « décennie du cerveau (en) » pour commémorer les avancées de la recherche sur le cerveau, et promouvoir les fonds pour les recherches futures[12],[13].
Structure
[modifier | modifier le code]
Le cerveau d'un homme adulte pèse près d'1,5 kg en moyenne (généralement de 1,3 à 1,4 kg, ce qui représente environ 2 % du poids corporel pour une personne de 60 kg)[14]. Il a un volume d'environ 1 130 cm3 chez la femme et 1 290 cm3 chez l'homme, bien qu'il existe des variations individuelles importantes[15]. Rapportés à une taille et une surface corporelle identique, les hommes ont en moyenne un cerveau 100 grammes plus lourd que celui des femmes[16], mais ces différences ne sont pas corrélées avec le quotient intellectuel ou d'autres mesures de performance cognitive[17].
Le cerveau humain est constitué d'environ 170 milliards de cellules dont 86 milliards de neurones en moyenne[18],[19], qui peuvent chacun former de 5 à 60 000 synapses[20]. La forte myélinisation des axones permet d'accélérer la vitesse de l'influx nerveux qui se propage de 1 m/s dans un axone amyélinisé à 100 m/s dans un axone myélinisé[21].
Le cerveau humain (partie la plus proéminente de l'encéphale), est situé au-dessus du cervelet et du tronc cérébral. Il comprend deux hémisphères cérébraux (formant avec des structures associées le télencéphale) et le diencéphale constitués des thalamus, hypothalamus, épithalamus et sous-thalamus.
L'encéphale humain possède la même structure que celui des vertébrés, avec certaines régions divergentes comparé à d'autres espèces : le cortex cérébral, l’amygdale, le thalamus et le cervelet[22]. La neuroanatomie le divise en six régions principales définies sur la base du développement du système nerveux à partir du tube neural : le télencéphale, le diencéphale, le mésencéphale, le cervelet, le pont, et le bulbe rachidien. Chacune de ces régions possède une structure interne complexe. Certaines régions du cerveau, comme le cortex cérébral ou le cervelet, sont formés de couches formant des replis sinueux, les circonvolutions cérébrales, qui permettent d'augmenter la surface corticale tout en logeant dans la boîte crânienne. Les autres régions du cerveau représentent des groupes de nombreux noyaux. Si des distinctions claires peuvent être établies à partir de la structure neurale, la chimie et la connectivité, des milliers de régions distinctes peuvent être identifiées dans le cerveau.
Structure et organisation du cortex cérébral
[modifier | modifier le code]

| Aire motrice primaire | |
| Aire motrice supplémentaire préfrontale | |
| Aire sensorielle primaire | |
| Aire d'association sensorielle | |
| Aire visuelle primaire | |
| Aire auditive primaire |
L'épaisseur du cortex cérébral est comprise entre 1 et 4,5 millimètres et sa surface avoisine deux mille centimètres carrés[23].
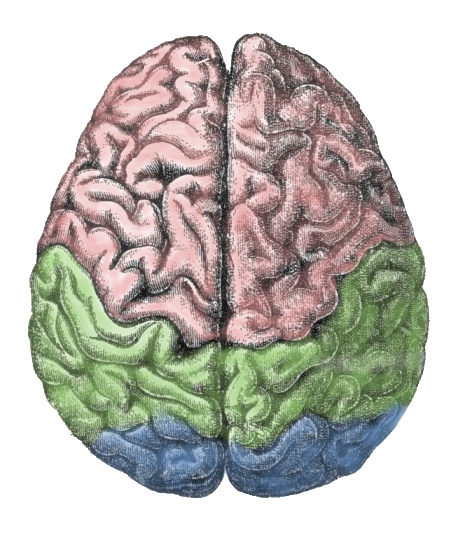
Il y aurait une évidente difficulté à loger toute cette surface dans la boîte crânienne. En fait le cortex est plissé par des sillons (sulcus en latin scientifique) ou scissures, sinueux et de profondeur variable, délimitant des crêtes appelées gyrus[24] ou circonvolutions cérébrales. Les scissures les plus profondes divisent le cortex en lobes. Selon leur situation, on parle de lobe frontal, pariétal, occipital et temporal.
Sous le cortex se trouve la substance blanche composée d'axones qui établissent les connexions entre corps cellulaires du cortex et d'autres parties du cerveau.
Le cortex cérébral est divisé en zones fonctionnelles, appelées aires (les trois grands types de zones étant les aires sensorielles, les aires motrices et les aires d'association), chacune assurant une fonction cognitive précise. Ce sont les études de Paul Broca en 1861 qui ont pour la première fois suggéré l'existence d'une telle organisation du néocortex[25]. Ces zones sont à peu près identiques pour tous les individus d'une même espèce, mais présentent de petites différences. Cette spécialisation est floue chez les mammifères primitifs et se précise au fur et à mesure que l'on se rapproche de l'humain.
Des recherches, dont les résultats furent publiés en 2016, ont conduit des chercheurs à délimiter 180 zones par hémisphère cérébral délimitées par des changements brusques de l'architecture corticale, la fonction, la connectivité, voire de la topographie. Les images ont été obtenues en utilisant la résonance magnétique multi-modales issues du projet connectome humain (en)[26],[N 1].
Différences entre le cerveau de la femme et celui de l'homme
[modifier | modifier le code]La plus grande différence structurelle entre le cerveau de l'homme et celui de la femme est la taille. Le cerveau de l'homme est plus gros (en moyenne 1 290 cm3 chez l'homme contre 1 130 cm3 chez la femme)[15] et plus lourd que celui de la femme de 10 à 15 %, cette donnée scientifique ayant « conduit plusieurs scientifiques du XIXe siècle à établir un lien entre la taille et le poids du cerveau d'une part et l'intelligence d'autre part »[27],[28].
Mais cette différence de taille du cerveau ne s'accompagne pas de différences de l'intelligence telle qu'elle est appréciée par la mesure globale du quotient intellectuel[29].
La littérature scientifique suggère « des différences de structure et aussi de fonction[Lesquelles ?] »[30], « qu'il existe bien des patterns d'activation différents en fonction du sexe pour des tâches variées comme la rotation mentale, le traitement verbal, la compréhension d'idiomes etc. Toutefois, ces résultats sont variables voire divergents d'une étude à l'autre et il n’y a pas de parallélisme strict entre les différences d'activation et les différences de performance »[31]. Si des différences anatomiques (notamment au niveau de l'amygdale, de l'hippocampe, du planum temporale et de l'insula)[32] et des différences jusqu'au niveau cellulaire et moléculaire, sont bien attestées, la complexité des interactions fonctionnelles rend difficile la corrélation de différences anatomiques avec des différences cognitives[15].
Selon une étude en 2014, les cerveaux masculins auraient davantage de connexions internes aux hémisphères, tandis que les cerveaux féminins auraient une meilleure connectivité entre les deux hémisphères[33]. Les auteurs en déduisent une meilleure prédisposition statistique des hommes en termes de coordination entre perception et action et inversement chez les femmes une meilleure prédisposition à combiner raisonnements analytiques et analogique. Cette étude a toutefois été critiquée pour mettre en avant des différences qui restent relatives sans les quantifier[34] et ainsi enfermer hommes et femmes dans des stéréotypes sexistes, opposant la femme multitâche à l'homme monotâche[35]. Ces stéréotypes sont largement repris dans des publications telles que Les hommes viennent de Mars, les femmes viennent de Vénus (clichés selon lesquels les hommes seraient plus agressifs et plus aptes à se déplacer ou à lire des cartes routières, les femmes plus dans l'empathie ou plus enclines aux compétences langagières)[36].
Une étude longitudinale basée sur l'imagerie médicale met en avant un décalage dans le temps du développement du cerveau entre hommes et femmes[37].
La neurobiologiste Catherine Vidal[38] dénonce le « neurosexisme » de ces stéréotypes à travers de nombreuses études qui voudraient montrer des différences intrinsèques entre cerveaux masculins et féminins. Elle privilégie les facteurs environnementaux aux facteurs innés, considérant que ces différences sont principalement dues à l'éducation et à la capacité du cerveau à se modifier en fonction de ce qui nous est appris (plasticité cérébrale)[39]. A contrario, les chercheurs Franck Ramus et Nicolas Gauvrit considèrent que la synthèse que fait Catherine Vidal des recherches scientifiques portant sur le cerveau et sur les différences entre les sexes « est extrêmement biaisée, incomplète, et que les arguments qu’elle utilise ne viennent pas à l’appui de ses conclusions »[40].
L'homme a plus de liquide cérébrospinal et de substance blanche (axones), et la femme a plus de substance grise (tissu neuronal et dendrites)[41].
Développement
[modifier | modifier le code]Le développement cérébral s'effectue selon plusieurs phases qui peuvent se chevaucher[42] :
- Neurogenèse : les neurones du cerveau humain commencent à se former autour de la 8e semaine de grossesse et sont pratiquement tous constitués à la 28e semaine ;
- Migration neuronale : la migration radiaire et tangentielle des neurones, de leur lieu de production à leur destination finale s'effectue sur de courtes (quelques mm) ou de longues distances (plusieurs cm) ;
- Synaptogenèse : elle débute dès la seconde moitié de la grossesse et culmine durant les premières années de vie. Elle est suivie de l'élagage synaptique, notamment à l'adolescence, qui consiste en la réduction de moitié du nombre des synapses. Le cortex cérébral est considéré comme pleinement mature vers 30 ans ;
- Apoptose neuronale : 15 à 50 % des neurones sont éliminés ;
- Myélinisation des aires primaires puis des aires associatives qui sont câblées et réorganisées en fonction des activités du nourrisson (toucher dont le développement est fonctionnel au bout de 11 semaines de grossesse, le sens de l'équilibre au bout de 21 semaines, l'odorat[43] et le sens gustatif au bout de 24 semaines, l'audition au bout de 32 semaines et enfin la vision)[20].
Fonctions
[modifier | modifier le code]La fonction principale du cerveau est de contrôler les actions de l'organisme à partir des informations sensorielles qui lui parviennent[44]. Les signaux sensoriels peuvent stimuler une réponse immédiate, moduler un schéma d'activité en cours, ou être emmagasinés pour un besoin futur. Ainsi, par le rôle central qu'il exerce dans la captation des stimuli externes, le cerveau occupe le rôle central dans la création de réponses à l'environnement. Le cerveau a aussi un rôle dans la régulation hormonale.
Le cerveau des chordés reçoit des signaux par les nerfs afférents de la part des différentes régions de l'organisme. Le cerveau interprète ces signaux et en tire une réponse fondée sur l'intégration des signaux électriques reçus, puis la transmet. Ce jeu de réception, d'intégration, et d'émission de signaux représente la fonction majeure du cerveau, qui explique à la fois les sensations, le mouvement, la mémoire et, on le suppose, la conscience.
Pour mener à bien sa complexe tâche, le cerveau est organisé en sous-systèmes fonctionnels, c'est-à-dire que certaines régions cérébrales traitent plus spécifiquement certains aspects de l'information. Cette division fonctionnelle n'est pas stricte et ces sous-systèmes peuvent être catégorisés de plusieurs façons : anatomiquement, chimiquement ou fonctionnellement. Une de ces catégorisations repose sur les neurotransmetteurs chimiques utilisés par les neurones pour communiquer. Une autre se base sur la manière dont chaque zone du cerveau contribue au traitement de l'information : les zones sensorielles amènent l'information au cerveau ; les signaux moteurs envoient l'information du cerveau jusqu'aux muscles et aux glandes ; les systèmes excitateurs modulent l'activité du cerveau en fonction du moment de la journée et de divers facteurs.
Le cerveau utilise principalement le glucose comme substrat énergétique et une perte de conscience peut survenir s'il en manque. La consommation énergétique du cerveau n'est pas particulièrement variable, mais les régions actives du cortex consomment plus d'énergie que les inactives.
Systèmes de neurotransmissions
[modifier | modifier le code]Selon le principe de Dale, chaque neurone du cerveau libère constamment le même neurotransmetteur chimique, ou la même combinaison de neurotransmetteurs, pour toutes les connexions synaptiques qu'il entretient avec d'autres neurones[45]. Un neurone peut donc être caractérisé en fonction des neurotransmetteurs qu'il libère bien qu'il existe quelques exceptions à ce principe. Les deux neurotransmetteurs les plus fréquents sont le glutamate, qui correspond généralement à un signal excitatoire, et l'acide γ-aminobutyrique (GABA), généralement inhibitoire. Les neurones utilisant ces deux neurotransmetteurs se retrouvent dans presque toutes les régions du cerveau et forment un large pourcentage des synapses du cerveau[46].
Les autres neurotransmetteurs, comme la sérotonine ou la noradrénaline, proviennent de neurones localisés dans des zones particulières du cerveau. D'autres neurotransmetteurs, comme l'acétylcholine ou la dopamine, proviennent de plusieurs endroits du cerveau, mais ne sont pas distribués de façon aussi ubiquitaire que le glutamate et le GABA. La grande majorité des drogues psychotropes agissent en altérant les systèmes de neurotransmetteurs qui ne sont pas directement impliqués dans les transmissions glutamatergiques ou GABAergiques[47].
Systèmes sensoriels
[modifier | modifier le code]Une fonction importante du cerveau est de traiter l'information reçue par les récepteurs sensoriels[48]. Contrairement aux idées reçues, les sens que peut capter le cerveau ne sont pas limités à cinq. Outre la vue, l'ouïe, le toucher, l'odorat, et le goût, le cerveau peut recevoir d'autres informations sensorielles comme la température, l'équilibre, la position des membres (proprioception), ou la composition chimique du sang. Toutes ces variables sont détectées par des récepteurs spécialisés qui transmettent les signaux vers le cerveau. Certaines espèces peuvent détecter des sens supplémentaires, comme la vision infrarouge des serpents, ou utiliser les sens « standards » de manière non conventionnelle, comme l'écholocation du système auditif des chauves-souris.
Chaque système sensoriel possède ses propres cellules sensorielles réceptrices. Ces cellules sont des neurones mais, contrairement à la majorité des neurones, ceux-ci ne sont pas contrôlés par les signaux synaptiques d'autres neurones. Au lieu de cela, ces cellules sensorielles possèdent des récepteurs membranaires qui sont stimulées par un facteur physique spécifique comme la lumière, la température, ou la pression. Les signaux de ces cellules sensorielles réceptrices parviennent jusqu'à la moelle épinière ou le cerveau par les nerfs afférents.
Pour la plupart des sens, il y a un noyau sensitif principal dans le tronc cérébral, ou un ensemble de noyaux, qui reçoit et réunit les signaux des cellules sensorielles réceptrices. Dans de nombreux cas, des zones secondaires sous-corticales se chargent d'extraire et de trier l'information. Chaque système sensoriel a également une région du thalamus qui lui est dédié et qui relaie l'information au cortex.
Pour chaque système sensoriel, une zone corticale primaire reçoit directement les signaux en provenance du relai thalamique. Habituellement, un groupe spécifique de zones corticales supérieures analyse également le signal sensoriel. Enfin, des zones multimodales du cortex combinent les signaux en provenance de différents systèmes sensoriels. À ce niveau, les signaux qui atteignent ces régions du cerveau sont considérés comme des signaux intégrés plutôt que comme des signaux strictement sensoriels[49].
Toutes ces étapes ont leurs exceptions. Ainsi, pour le toucher, les signaux sensoriels sont principalement reçus au niveau de la moelle épinière, au niveau de neurones qui projettent ensuite l'information au tronc cérébral[50]. Pour l'odorat, il n'y a pas de relai dans le thalamus, le signal est transmis directement de la zone primaire, le bulbe olfactif, vers le cortex[51].
Systèmes moteurs
[modifier | modifier le code]Les systèmes moteurs sont les zones du cerveau responsables directement ou indirectement des mouvements du corps, en agissant sur les muscles. À l'exception des muscles contrôlant les yeux, tous les muscles squelettiques de l'organisme sont directement innervés par des neurones moteurs de la moelle épinière. Ils sont donc le dernier maillon de la chaîne du système psychomoteur[52]. Les neurones moteurs spinaux sont contrôlés à la fois par des circuits neuronaux propres à la moelle épinière, et par des influx efférents du cerveau. Les circuits spinaux intrinsèques hébergent plusieurs réactions réflexes, ainsi que certains schémas de mouvements comme les mouvements rythmiques tels que la marche ou la nage[53]. Les connexions efférentes du cerveau permettent quant à elles, des contrôles plus sophistiqués.
Un certain nombre de zones du cerveau sont connectées directement à la moelle épinière[54]. Au niveau le plus bas se trouvent les zones motrices situées dans le bulbe rachidien et le pont. Au-dessus se situent les zones du mésencéphale, comme le noyau rouge, qui sont responsables de la coordination des mouvements. À un niveau supérieur se trouve le cortex moteur primaire, une bande de tissu cérébral localisée à la lisière postérieure du lobe frontal. Le cortex moteur primaire transmet ses commandes motrices aux zones motrices sous-corticales, mais également directement à la moelle épinière par le biais du faisceau pyramidal. Les influx nerveux de ce faisceau cortico-spinal transmettent les mouvements fins volontaires. D'autres zones motrices du cerveau ne sont pas directement reliées à la moelle épinière, mais agissent sur les zones motrices primaires corticales ou sous-corticales. Quelques-unes de ces zones secondaires les plus importantes sont le cortex prémoteur, impliqués dans la coordination des mouvements de différentes parties du corps, les ganglions de la base, dont la fonction principale semble être la sélection de l'action, et le cervelet, qui module et optimise les informations pour rendre les mouvements plus précis.
Le cerveau et la moelle épinière contiennent également un réseau neuronal qui contrôle le système nerveux autonome, la partie du système nerveux responsable des fonctions automatiques. Non soumis au contrôle volontaire, le système nerveux autonome contrôle notamment la régulation hormonale et l'activité des muscles lisses et du muscle cardiaque. Le système nerveux autonome agit à différents niveaux comme le rythme cardiaque, la digestion, la respiration, la salivation, la miction, la sueur ou l'excitation sexuelle.
Systèmes d'éveil
[modifier | modifier le code]Un des aspects les plus visibles du comportement animal est le cycle journalier veille-sommeil-rêve. L'éveil et l'attention sont aussi modulés à une échelle de temps plus fine, par un réseau de zones cérébrales[55].
Un composant clé du système d'éveil est le noyau suprachiasmatique, petite région de l'hypothalamus localisée directement au-dessus du point de croisement des nerfs optiques[56]. Le noyau suprachiasmatique renferme l'horloge biologique centrale de l'organisme. Les neurones de ce noyau montrent un niveau d'activité qui augmente ou diminue sur une période d'environ 24 heures, le rythme circadien : cette activité fluctuante est dirigée par des changements rythmiques exprimés par un groupe de gènes horlogers. Le noyau suprachiasmatique reçoit généralement des signaux en provenance des nerfs optiques qui permettent de calibrer l'horloge biologique à partir des cycles jour-nuit.
Le noyau suprachiasmatique se projette dans un ensemble de zones cérébrales (situées au niveau de l'hypothalamus et du tronc cérébral) qui sont impliqués dans la mise en œuvre des cycles jour-nuit. Un composant important du système est la formation réticulée, un groupe d'amas neuronaux s'étendant dans le tronc cérébral[55]. Les neurones réticulés envoient des signaux vers le thalamus, qui répond en envoyant des signaux à différentes régions du cortex qui régule le niveau d'activité.
Le sommeil implique de profondes modifications dans l'activité cérébrale[57]. Le cerveau ne s'éteint pas pendant le sommeil, l'activité cérébrale se poursuit mais est modifiée. En fait, il existe deux types de sommeil : le sommeil paradoxal (avec rêves) et le sommeil non paradoxal (généralement sans rêves). Ces deux sommeils se répètent selon un schéma légèrement différent à chaque sommeil. Trois grands types de schéma d'activité cérébrale peuvent être distingués : sommeil paradoxal, sommeil léger, et sommeil profond. Pendant le sommeil profond, l'activité du cortex prend la forme de larges ondes synchronisées tandis que ces ondes sont désynchronisées pendant l'état de rêve. Les niveaux de noradrénaline et de sérotonine tombent au cours du sommeil profond, et approchent du niveau zéro pendant le sommeil paradoxal, tandis que les niveaux d'acétylcholine présentent un schéma inverse.
Le cycle du sommeil se divise en 5 stades. Au stade 1, le sommeil lent apparaît lorsque l'on s'allonge et ferme les yeux, des rêves courts ou de brèves pensées sont parfois rapportés durant ce stade qui dure de 3 à 12 minutes. La seconde phase du sommeil lent est un sommeil léger, c'est la phase la plus longue occupant près de 50 % du temps de sommeil d'une nuit. Le stade 3 est le passage du sommeil moyennement profond à profond, les muscles ont encore du tonus mais on est très peu réactif aux stimulations extérieures. Au niveau 4, nous sommes au plus profond de notre sommeil, l'activité neuronale est à son plus bas, la température du cerveau est également basse, la respiration, le rythme cardiaque et la pression sanguine sont ralentis. Le stade 5 est le dernier, le sommeil est paradoxal, l'activité électrique du cerveau est très importante, les yeux bougent rapidement, bien que le reste du corps se trouve en état d'atonie musculaire. Un cycle du sommeil dure de 70 à 90 minutes et se reproduit 4 à 6 fois en une nuit[58].
Latéralisation et dominance cérébrale
[modifier | modifier le code]Évolution
[modifier | modifier le code]Évolution de la capacité crânienne
[modifier | modifier le code]Au cours de l'évolution des Hominina, le volume du cerveau humain est passé d'environ 600 cm3 chez l'Homo habilis à environ 1 500 cm3 chez l'Homo neanderthalensis. On observe ensuite une diminution au cours des 28 000 dernières années, le cerveau masculin passant de 1 500 cm3 à 1 350 cm3, et le cerveau féminin diminuant dans la même proportion[59]. À titre de comparaison, l'Homo erectus, un parent de l'humain moderne, avait un cerveau de 1 100 cm3. Cependant le petit Homo floresiensis, doté d'un cerveau de seulement 380 cm3, donc trois fois moins volumineux que celui de l'Homo erectus, connaissait l'usage du feu, chassait et fabriquait des outils de pierre au moins aussi élaborés que ceux de l'Homo erectus[60]. Si la différence de taille entre les cerveaux de Homo sapiens et Homo Neanerthalis était faible, l'assignation des capacités était différente, avec une plus grande place accordée à la vision et au mouvement chez l'Homme de Neanderthal, tandis que chez Homo sapiens, les activités de cognition liée aux réseaux sociaux étaient plus développées[61].
Cette évolution de la taille a été précédée de la réorganisation des régions corticales[62].
Les principaux facteurs qui auraient favorisé cet accroissement de volume font l'objet de débats au sein des chercheurs. Ils proposent notamment l'incorporation de la viande à la prise alimentaire des hominidés et généralisation de la cuisson, ce qui entraîne une réduction du même ordre de l'intestin[N 2] coûteux en énergie[63] ; la flexibilité cognitive et apprentissage qui auraient permis aux hominidés d'exploiter des ressources éphémères (plantes, arbres fruitiers) et des relations plus complexes dans les sociétés fission-fusion[64] ; le développement des capacités cognitives des hominidés à l'époque où ils étaient des proies (nécessité de s’organiser, de monter la garde la nuit, d'apprendre à communiquer, pour échapper aux prédateurs)[65] ; le rôle des stratégies cynégétiques complexes et collectives et des rituels très hiérarchisés qui auraient favorisé l'émergence de l'homme sociétal et l'accroissement de volume[66].
Différences dans l'organisation du cortex cérébral avec les autres mammifères
[modifier | modifier le code]Le cortex cérébral est composé de neurones pyramidaux (excitateurs) et d'interneurones inhibiteurs. Le cortex cérébral humain contient les mêmes types de neurones que les autres espèces mammifères, avec toutefois des différences quantitatives : l'humain est l'une des espèces (avec les autres primates et les cétacés) avec le plus de neurones corticaux (nombre absolu et nombre rapporté au poids de l'individu), et, comme les autres primates, contient une plus grande proportion de certains types cellulaires : le cortex cérébral humain contient davantage de neurones pyramidaux des couches II/III du néocortex (impliquées dans les connexions entre régions du cortex), plus d'interneurones inhibiteurs (20-25%) ainsi que plus de cellules gliales[67],[68].
Ces modifications de composition cellulaire sont en grande partie dues à un développement (corticogenèse) plus long chez l'humain comparé aux autres espèces (environ 3 mois chez l'humain comparé à une semaine chez la souris par exemple) ainsi qu'à une plus grande diversité de cellules progénitrices[67].
À l'échelle cellulaire, les neurones pyramidaux humains présentent une plus grande complexité dans leur morphologie (arbre dendritique) ainsi qu'une plus grande densité d'épines dendritiques, structures post-synaptiques impliquées dans la transmission nerveuse[67]
La taille du cerveau ainsi que l'augmentation du nombre de neurones corticaux et de certains types de cellules neurales ne peut expliquer à lui seul l'évolution du cerveau chez l'humain. La maturation des réseaux neuronaux est aussi en jeu, notamment dans sa temporalité. Le développement neuronal est beaucoup plus lent chez l'humain, reste immature plus longtemps : la néoténie. Cela permet aux réseaux neuronaux de rester "plastiques" sur une plus longue période et ainsi d'augmenter les capacités d'apprentissage. Cette propriété est intrinsèque aux neurones pyramidaux : par exemple, la transplantation, dans un cerveau de souris, de neurones différenciés à partir de cellules pluripotentes humaines, de chimpanzé, de macaque ou de souris montre un gradient de vitesse de développement, du plus vite chez la souris (~1 mois pour être mature) au plus lent chez l'humain (~9 mois pour être mature)[67]. Cette propriété est intrinsèque aux neurones humains[69].
Au-delà du développement des neurones du cortex cérébral, leurs propriétés fonctionnelles à l'âge adulte divergent elles-aussi comparé aux autres animaux dont les autres primates (macaques, marmousets)[70]. Les neurones pyramidaux humains sont plus grands, complexes et forment plus de synapses que les neurones d'autres espèces[70]. De plus, les neurones pyramidaux humains sont électriquement moins excitables que ceux des autres espèces[71]. Cela pourrait être dû à la plus grande taille des neurones humains[71], une plus grande expression de certains canaux ioniques (HCN) dans les dendrites des neurones[72], des changements de propriétés des canaux ioniques à la zone de gâchette des neurones (qui génère les potentiels d'action)[73], ou encore une plus grande inhibition de ces canaux ioniques à la zone de gâchette[74]. Les conséquences évolutives de cette plus faible excitabilité neuronale sont encore inconnues. Elle pourrait permettre un encodage d'informations plus complexe et précis dans le temps, ce qui est par exemple critique dans le langage. En effet, le cortex auditif, impliqué dans le langage, met plus de temps à traiter une information auditive comparé au chimpanzé et au macaque, ce qui pourrait lui permettre de traiter des informations auditives plus complexes ; une plus faible excitabilité pourrait permettre cela[75].
Métabolisme
[modifier | modifier le code]Le cerveau ne représente que 2 % de la masse du corps humain, mais 20 à 25 % de sa consommation énergétique[76].
Une étude de 2016 montre une corrélation chez la souris, le singe et l'humain entre le niveau sanguin d'une enzyme, la myokine cathepsine B (CTSB) avec la forme physique et la mémoire. Chez la souris, il a été démontré que l'augmentation de la production de cathepsine B améliore les niveaux de neurotrophine dans les cellules souches adultes hippocampiques pour conduire à l'amélioration des performances de la mémoire. Cette protéine est produite par les muscles lors de l'activité physique. Si le gène de la CTSB est désactivé chez la souris, l'activité physique ne produit plus aucune amélioration pour la mémoire. Cette étude suggère donc que la CTSB pourrait stimuler la formation de nouveaux neurones de l'hippocampe dans le cerveau humain[77].
Pathologies
[modifier | modifier le code]Trouble du spectre de l'alcoolisation fœtale
[modifier | modifier le code]La construction du cerveau pendant la période embryonnaire ou fœtale peut être affectée si la mère consomme de l'alcool. Les séquelles sont permanentes et peuvent causer un trouble du spectre de l'alcoolisation fœtale.
Surdité centrale
[modifier | modifier le code]Également appelée surdité cognitive, ou agnosie auditive, la surdité centrale est provoquée par des troubles dont l'origine se situe dans les aires auditives du cerveau, et non au niveau du système nerveux périphérique (dans l'oreille interne ou au niveau du nerf auditif).
Notes et références
[modifier | modifier le code]Notes
[modifier | modifier le code]- L'étude a porté sur 210 jeunes adultes en bonne santé.
- Chez la plupart des mammifères herbivores de la même masse corporelle que l'humain, le cerveau est environ cinq fois plus petit que le cerveau humain, tandis que l'intestin est deux fois plus volumineux car il est adapté à une alimentation en partie non digestible (fibres végétales) grâce au microbiote intestinal (bactéries équipées des enzymes pouvant digérer la cellulose).
Références
[modifier | modifier le code]- André Bourguignon, Cyrille Koupernik, Pierre-Marie Lledo, Bernard Mazoyer, Jean-Didier Vincent, « CERVEAU HUMAIN », sur universalis.fr (consulté le )
- Gérald Fournier, Évolution et civilisation : de l'anthropologie de Charles Darwin à l'économie évolutionniste étendue, Fournier, , 815 p. (ISBN 978-2-9540304-0-1), p. 484.
- Xavier Seron et Marc Jeannerod, Neuropsychologie humaine, Éditions Mardaga, , p. 153.
- (en) S. Finger, Origins of neuroscience : A history of explorations into brain function, Oxford University Press, , 480 p. (ISBN 978-0-19-514694-3, présentation en ligne), p. 14
- « De la psychobiologie aux neurosciences cognitives »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?) [PDF] (consulté le ).
- (en) R. Hendrickson, The facts on file encyclopedia of word and phrase origins, New York, Facts on File, , 4e éd., 948 p. (ISBN 978-0-8160-6966-8, présentation en ligne).
- « Modèles cognitifs de la mémoire » [PDF], sur applis.univ-tours.fr.
- Bernard Mazoyer et John W. Belliveau, « Les nouveaux progrès de l'imagerie », sur larecherche.fr, .
- (en) F. E. Bloom et J. P. Swazey (dir.), The Neurosciences, paths of discovery, vol. 1973, MIT Press, , 622 p. (ISBN 978-0-262-23072-8, présentation en ligne), p. 211.
- (en) Louis Sokoloff, « The metabolism of the central nervous system in vivo », Neurophysiology, vol. 3, , p. 1843-1864.
- (en) M. Piccolino, « Fifty years of the Hodgkin-Huxley era », Trends Neurosci., vol. 25, no 11, , p. 552-553 (ISSN 0166-2236, PMID 12392928, DOI 10.1016/S0166-2236(02)02276-2).
- (en) George H. W. Bush, « Project on the Decade of the Brain », sur loc.gov, (consulté le ).
- (en) E. G. Jones et L. M. Mendell, « Assessing the decade of the brain », Science, vol. 284, no 5415, , p. 739 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.284.5415.739).
- Denis Le Bihan, Le cerveau de cristal. Ce que nous révèle la neuro-imagerie, Odile Jacob, .
- (en) Cosgrove, KP, Mazure CM, Staley JK, « Evolving knowledge of sex differences in brain structure, function, and chemistry », Biol Psychiat, vol. 62, no 8, , p. 847-855.
- (en) C. Davison Ankney, « Sex differences in relative brain size : The mismeasure of woman, too ? », Intelligence, vol. 16, nos 3–4, , p. 329-336 (DOI 10.1016/0160-2896(92)90013-H).
- (en) RC Gur, BI Turetsky, M Matsui, M Yan, W Bilker, P Hughett et RE Gur, « Sex differences in brain gray and white matter in healthy young adults : correlations with cognitive performance », The Journal of Neuroscience, vol. 19, no 10, , p. 4065-4072 (lire en ligne).
- (en) Frederico A. C. Azevedo et al., « Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain », The Journal of Comparative Neurology, vol. 513, no 5, (DOI 10.1002/cne.21974, lire en ligne).
- (en) Suzana Herculano-Houzel, « The human brain in numbers: a linearly scaled-up primate brain », Frontiers in Human Neuroscience, vol. 3, (ISSN 1662-5161, DOI 10.3389/neuro.09.031.2009, lire en ligne).
- Bernard Sablonnière, Le cerveau. Les clés de son développement et de sa longévité, Jean-Claude Gawsewitch Éditeur, (ISBN 978-2-35013-445-1), p. 256.
- Karen Huffman, Introduction à la psychologie, De Boeck Supérieur, , p. 84.
- (en) Luiz Pessoa, « Understanding brain networks and brain organization », Physics of Life Reviews, vol. 11, no 3, , p. 400-435 (PMID 24819881, PMCID PMC4157099, DOI 10.1016/j.plrev.2014.03.005, lire en ligne, consulté le )
- (en) B. Fischl et A. M. Dale, « Measuring the thickness of the human cerebral cortex from magnetic resonance images », Proc Natl Acad Sci U S A, vol. 20, no 97, , p. 11050-11055.
- Le terme de latin anatomique gyrus a pour pluriel gyri, en terminologie française le pluriel est gyrus.
- R. Saban, Conception de la physiologie cérébrale de François-Joseph Gall à Paul Broca : les localisations des fonctions cérébrales, in Biométrie humaine et anthropologie - Tome 20, numéros 3-4, juillet - décembre 2002, p. 195-203. « http://edition.cens.cnrs.fr/revue/bha/2002/v20/n3-4/011747ar.html »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?), (Cf. marque 32).
- « A multi-modal parcellation of human cerebral cortex », Matthew F. Glasser et col., Nature, vol. 536, p. 171-178, 11 août 2016, doi:10.1038/nature18933.
- Cécile Guillaume, « Le cerveau a-t-il un sexe ? », Les Cahiers Dynamiques, vol. 1, no 58, , p. 33.
- Émile Durkheim, « De la division du travail, livre 1 », (consulté le ) : « On dirait que les deux grandes fonctions de la vie psychique se sont comme dissociées, que l'un des sexes a accaparé les fonctions affectives et l'autre les fonctions intellectuelles. (…) cet écart progressif serait dû, à la fois, au développement considérable des crânes masculins et à un stationnement ou même une régression des crânes féminins. », p. 64-65
- (en) Ulric Neisser et coll., « Intelligence: Knowns and unknowns », American Psychologist, vol. 51, no 2, , p. 77-101 (DOI 10.1037/0003-066X.51.2.77).
- Mariana Alonso, Le sexe du cerveau in La Recherche juillet-août 2019 https://www.larecherche.fr/chronique-neurosciences-id%C3%A9es/le-sexe-du-cerveau
- Cécile Guillaume, op. cit., p. 34
- (en) Amber N.V. Ruigrok, Gholamreza Salimi-Khorshidi, Meng-Chuan Lai, Simon Baron-Cohen, Michael V.Lombardo, Roger J. Tait, John Suckling, « A meta-analysis of sex differences in human brain structure », Neuroscience & Biobehavioral Reviews, vol. 39, , p. 34-50 (DOI 10.1016/j.neubiorev.2013.12.004).
- (en) Madhura Ingalhalikar, Alex Smith, Drew Parker, Theodore D. Satterthwaite, Mark A. Elliott, Kosha Rupare, Hakon Hakonarson, Raquel E. Gur, Ruben C. Gur, & Ragini Verma, « Sex differences in the structural connectome of the human brain », Proc Natl Acad Sci USA, vol. 11, no 2, , p. 823-828 (DOI 10.1073/pnas.1316909110).
- (en) Daphna Joel and Ricardo Tarrasch, On the mis-presentation and misinterpretation of gender-related data: The case of Ingalhalikar’s human connectome study, PNAS, 11 février 2014
- Sabine Casalonga, « Cerveau de Vénus, cerveau de Mars. Quelles différences ? », Le Monde de l'intelligence, no 35, , p. 17.
- Pascal Picq, Philippe Brenot, Le Sexe, l’Homme et l’Évolution, Odile Jacob, , p. 159.
- Lenroot et al., 2007 DOI 10.1016/j.neuroimage.2007.03.053
- Catherine Vidal, Nos cerveaux, tous pareils, tous différents !, Belin, 2015.
- Martine Fournier, Masculin : Féminin. Pluriel, Sciences Humaines Éditions, , p. 94.
- Nicolas Gauvrit et Frank Ramus, « La « méthode Vidal » », Science et pseudo-sciences, no 309, (lire en ligne).
- « Taille du cerveau : comparaison femme-homme », sur futura-sciences.com (consulté le )
- (en) GZ Tau et BS Peterson, « Normal Development of Brain Circuits », Neuropsychopharmacology, vol. 35, no 1, , p. 147–168 (DOI 10.1038/npp.2009.115).
- Benoist Schaal. Le fœtus perçoit les odeurs et s'en souvient : la formation d'attentes sensorielles et leurs implications. Médecine & Enfance, 2010, 30 (Hors Série "Aspects Sensoriels de l'Alimentation de l'Enfant"), pp.20-34.
- (en) T. J. Carew, Behavioral neurobiology : The cellular organization of natural behavior, Sinauer Associates, , 435 p. (ISBN 978-0-87893-092-0, présentation en ligne), chap. 1
- Kandel 2000, ch. 15
- (en) P. L. McGeer et E. G. McGeer, Basic neurochemistry : Molecular, cellular and medical aspects, New York, Raven Press, , 4e éd., 984 p. (ISBN 978-0-88167-343-2, présentation en ligne), chap. 15 (« Amino acid neurotransmitters »), p. 311-332
- (en) J. R. Cooper, F. E. Bloom et R. H. Roth, The biochemical basis of neuropharmacology, Oxford University Press, , 8e éd., 405 p. (ISBN 978-0-19-514008-8, présentation en ligne)
- Kandel 2000, ch. 21
- Kandel 2000, ch. 21 et 30
- Kandel 2000, ch. 23
- Kandel 2000, ch. 32
- Kandel 2000, ch. 34
- Kandel 2000, ch. 36-37
- Kandel 2000, ch. 33
- Kandel 2000, ch. 45
- (en) M. C. Antle et R. Silver, « Orchestrating time: arrangements of the brain circadian clock », Trends in Neurosciences, vol. 28, no 3, , p. 145-151 (ISSN 0166-2236, PMID 15749168, DOI 10.1016/j.tins.2005.01.003, lire en ligne)
- Kandel 2000, ch. 47
- « Les cycles du sommeil » (consulté le )
- (en) « If Modern Humans Are So Smart, Why Are Our Brains Shrinking? », DiscoverMagazine.com, (consulté le ).
- (en) P Brown, T Sutikna, MJ Morwood et al., « A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia », Nature, vol. 431, no 7012, , p. 1055-1061 (PMID 15514638, DOI 10.1038/nature02999).
- (en) Jennifer Viegas, « Brain comparison suggests that Neanderthals lacked social skills », NBC News, (consulté le ).
- (en) Lauren A. Gonzales, Brenda R. Benefit, Monte L. McCrossin & Fred Spoor, « Cerebral complexity preceded enlarged brain size and reduced olfactory bulbs in Old World monkeys », Nature Communications, vol. 6, no 7580, (DOI 10.1038/ncomms8580).
- (en) Leslie C. Aiello & Peter Wheeler, « The Expensive-Tissue Hypothesis: The Brain and the Digestive System in Human and Primate Evolution », Current Anthropology, vol. 36, no 2, , p. 199-221.
- (en) R. Potts, « Paleoenvironmental basis of cognitive evolution in great apes », Am. J. Primatol, vol. 62, no 3, , p. 209–228.
- (en) Donna Hart, Robert Sussman, Man the Hunted : Primates, Predators, and Human Evolution, Westview Press Inc., , p. 12.
- Marylène Patou-Mathis, Mangeurs de viande : de la préhistoire à nos jours, Perrin, , p. 128.
- (en) Baptiste Libé-Philippot et Pierre Vanderhaeghen, « Cellular and Molecular Mechanisms Linking Human Cortical Development and Evolution », Annual Review of Genetics, vol. 55, no 1, , annurev–genet–071719-020705 (ISSN 0066-4197 et 1545-2948, DOI 10.1146/annurev-genet-071719-020705, lire en ligne, consulté le )
- (en) Jim Berg, Staci A. Sorensen, Jonathan T. Ting et Jeremy A. Miller, « Human neocortical expansion involves glutamatergic neuron diversification », Nature, vol. 598, no 7879, , p. 151–158 (ISSN 1476-4687, DOI 10.1038/s41586-021-03813-8, lire en ligne, consulté le )
- « Redirecting » (DOI 10.1016/j.neuron.2024.08.021, consulté le )
- (en) Pierre Vanderhaeghen et Franck Polleux, « Developmental mechanisms underlying the evolution of human cortical circuits », Nature Reviews Neuroscience, vol. 24, no 4, , p. 213–232 (ISSN 1471-0048, DOI 10.1038/s41583-023-00675-z, lire en ligne, consulté le )
- (en) Lou Beaulieu-Laroche, Norma J. Brown, Marissa Hansen et Enrique H. S. Toloza, « Allometric rules for mammalian cortical layer 5 neuron biophysics », Nature, vol. 600, no 7888, , p. 274–278 (ISSN 1476-4687, DOI 10.1038/s41586-021-04072-3, lire en ligne, consulté le )
- Brian E. Kalmbach, Anatoly Buchin, Brian Long et Jennie Close, « h-Channels Contribute to Divergent Intrinsic Membrane Properties of Supragranular Pyramidal Neurons in Human versus Mouse Cerebral Cortex », Neuron, vol. 100, no 5, , p. 1194-1208.e5 (ISSN 0896-6273, DOI 10.1016/j.neuron.2018.10.012, lire en ligne, consulté le )
- (en) René Wilbers, Verjinia D. Metodieva, Sarah Duverdin et Djai B. Heyer, « Human voltage-gated Na + and K + channel properties underlie sustained fast AP signaling », Science Advances, vol. 9, no 41, (ISSN 2375-2548, PMID 37824607, PMCID PMC10569700, DOI 10.1126/sciadv.ade3300, lire en ligne, consulté le )
- Baptiste Libé-Philippot, Amélie Lejeune, Keimpe Wierda et Nikolaos Louros, « LRRC37B is a human modifier of voltage-gated sodium channels and axon excitability in cortical neurons », Cell, vol. 186, no 26, , p. 5766-5783.e25 (ISSN 0092-8674, DOI 10.1016/j.cell.2023.11.028, lire en ligne, consulté le )
- (en) Kosuke Itoh, Naho Konoike, Masafumi Nejime et Haruhiko Iwaoki, « Cerebral cortical processing time is elongated in human brain evolution », Scientific Reports, vol. 12, no 1, , p. 1103 (ISSN 2045-2322, DOI 10.1038/s41598-022-05053-w, lire en ligne, consulté le )
- Pascal Picq, La plus belle histoire du langage, Paris, Seuil, , 184 p. (ISBN 978-2-02-040667-3), p. 69.
- « Running-Induced Systemic Cathepsin B Secretion Is Associated with Memory Function », Hyo Youl Moon et col., Cell Metabolism, Volume 24, no. 2, p. 332-340, 9 août 2016.
Voir aussi
[modifier | modifier le code]Bibliographie
[modifier | modifier le code]- Patrick Berche, Les Sortilèges du cerveau : L'histoire inédite de ce qui se passe dans nos têtes, Paris, Flammarion, , 448 p. (ISBN 978-2-08-129536-0, présentation en ligne).
- P. Buser et R. Lestienne, Cerveau, information, connaissance, CNRS Éditions, , 227 p. (ISBN 978-2-271-05773-0, présentation en ligne).
- J.-P. Changeux, Du vrai, du beau, du bien : Une nouvelle approche neuronale, Paris, Éditions Odile Jacob, , 544 p. (ISBN 978-2-7381-1904-9, présentation en ligne).
- A. R. Damasio (trad. de l'anglais), L'autre moi-même : Les nouvelles cartes du cerveau, de la conscience et des émotions, Paris, Éditions Odile Jacob, , 415 p. (ISBN 978-2-7381-1940-7, présentation en ligne).
- A. R. Damasio et M. Blanc, L'erreur de Descartes : La raison des émotions, Éditions Odile Jacob, , 368 p. (ISBN 978-2-7381-1713-7, présentation en ligne).
- Barbara Demeneix, Le Cerveau endommagé : Comment la pollution altère notre intelligence et notre santé, Paris, Odile Jacob, coll. « Sciences », , 416 p. (ISBN 9782738133915).
- J.-J. Feldmeyer, Le cerveau, Paris, Le Cavalier Bleu, coll. « Idées reçues » (no 131), , 127 p. (ISBN 978-2-84670-155-6, présentation en ligne).
- O. Houdé, B. Mazoyer, N. Tzourio-Mazoyer et F. Crivello, Cerveau et psychologie : Introduction à l'imagerie cérébrale anatomique et fonctionnelle, Paris, Presses universitaires de France, , 609 p. (ISBN 978-2-13-050912-7, présentation en ligne).
- M. Imbert, Traité du cerveau, Éditions Odile Jacob, , 532 p. (ISBN 978-2-7381-1665-9, présentation en ligne).
- J. E. LeDoux (trad. P. Kaldy), Le cerveau des émotions : Les mystérieux fondements de notre vie émotionnelle, Odile Jacob, , 374 p. (ISBN 978-2-7381-1633-8, présentation en ligne).
- M. Jeannerod, Le cerveau volontaire, Paris, Éditions Odile Jacob, , 297 p. (ISBN 978-2-7381-2357-2, présentation en ligne).
- R. Jouvent, Le cerveau magicien : De la réalité au plaisir psychique, Paris, Éditions Odile Jacob, , 252 p. (ISBN 978-2-7381-1879-0, présentation en ligne).
- B. Kolb et I. Q. Whishaw, Cerveau et comportement [« Neurosciences & cognition »], De Boeck Université, , 672 p. (ISBN 978-2-7445-0137-1, présentation en ligne).
- J.-D. Vincent, Voyage extraordinaire au centre du cerveau, Paris, Éditions Odile Jacob, , 455 p. (ISBN 978-2-7381-1935-3, présentation en ligne).
- J.-D. Vincent, Biologie des passions, Éditions Odile Jacob, , 341 p. (ISBN 978-2-7381-0737-4, présentation en ligne).
Articles connexes
[modifier | modifier le code]- Évolution du cerveau
- Système nerveux
- Crâne humain
- Human Brain Project
- Mythe de l'utilisation incomplète du cerveau
- Réseau du mode par défaut
Liens externes
[modifier | modifier le code]
- Ressources relatives à la santé :
- Notice dans un dictionnaire ou une encyclopédie généraliste :
- (en) IBRO (International Brain Research Organization)
- (en) Society for Neuroscience
- (en) Brain Research News from ScienceDaily
- (en) 3D Brain
- Le cerveau à tous les niveaux