Enzima limitada por difusión
Una enzima limitada por difusión es una enzima que cataliza una reacción química en forma tan eficiente, que el paso limitante de la velocidad es el de difusión; ya sea el de sustrato hacia el sitio activo, o el del producto hacia el medio.[2]
Esta propiedad también se denomina perfección cinética o perfección catalítica. Ya que la velocidad de catálisis de estas enzimas se encuentra determinada por una reacción limitada por difusión, representa una restricción física intrínseca a la evolución (un pico de máxima en el paisaje adaptativo); por lo que las enzimas perfectas limitadas por difusión son muy pocas. La mayor parte de las enzimas catalizan sus respectivas reacciones a velocidades que son entre 1000 y 10000 veces menores que ese límite. Esto es debido tanto a las limitaciones químicas de reacciones dificultosas, como a las limitaciones evolutivas debidas a que tan altas tasas de reacción no confieren una ventaja adaptativa.[1]
Historia
[editar]
La teoría de una reacción controlada por difusión fue utilizada originalmente por R.A. Alberty, Gordon Hammes, y Manfred Eigen para estimar el límite superior de una reacción de tipo enzima-sustrato.[3][4] De acuerdo con su estimación,[3][4] el límite superior para una reacción de tipo enzima-sustrato era de 109 M−1 s−1.
En 1972, se observó que en la deshidratación del H
2CO
3 catalizada por la anhidrasa carbónica, la constante catalítica de segundo orden obtenida experimentalmente estaba en el orden de 1,5 × 1010 M−1 s−1,[5] la cual es un orden de magnitud mayor que el límite estimado por Alberty, Hammes, y Eigen basados en un modelo simplificado.[3][4]
Para resolver esta aparente paradoja,[6] el profesor Kuo-Chen Chou y su equipo de colaboradores propusieron un modelo que tenía en cuenta el factor espacial, y el factor de campo de fuerzas entre la enzima y su sustrato, y encontraron que el límite superior podía alcanzar tasas de 1010 M−1 s−1,[7][8][9] y que podía ser utilizado para explicar algunas velocidades de reacción sorprendentemente altas observadas en biología molecular.[5][10][11]
El nuevo límite superior encontrado por Chou y colaboradores, para las reacciones enzima-sustrato fue posteriormente analizado y discutido por una serie de estudios subsecuentes.[12][13][14]
Una comparación detallada entre el modelo simplificado Alberty-Hammes-Eigen (a) y el modelo de Chou (b) para el cálculo de la velocidad de reacción de una enzima limitada por difusión con su sustrato, o el límite superior de una reacción de tipo enzima-sustrato, fue elaborada en la publicación de Zhou, del año 1982.[15]
Mecanismo
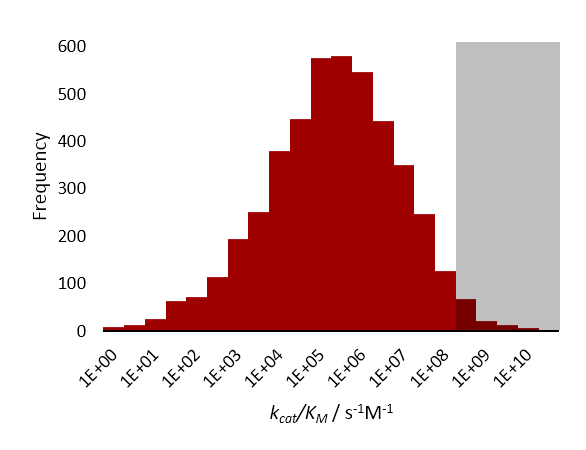
[editar]Las enzimas cinéticamente perfectas poseen una constante de especificidad, kcat/Km, en el oreden de 108 a 109 M−1 s−1. Donde la velocidad de la reacción catalizada por la enzima se encuentra limitada por difusión, y por lo tanto la enzima "procesa" al sustrato en su sitio activo, antes incluso de que otra molécula de sustrato pueda ingresar.[1])
Algunas enzimas funcionan con cinéticas que son más rápidas que las velocidades de difusión, lo cual parecería ser imposible. Existen varios mecanismos involucrados que pueden explicar este fenómeno. Se cree que algunas proteínas son capaces de acelerar el proceso de catálisis dirigiendo y preorientando a las moléculas de sustrato haciendo uso de campos eléctricos dipolares. Algunos hacen uso de efectos mecánico-cuánticos, tales como la explicación de efecto túnel de protones o electrones a través de las barreras de activación, aunque el efecto túnel de protones sigue siendo aún una idea controversial.[16][17]
Evolución
[editar]No es importante que no existan muchas enzimas cinéticamente perfectas. Esto puede ser explicado en términos de selección natural. Un aumento en la velocidad catalítica puede ser favorecido si confiere alguna ventaja para el organismo. Sin embargo, cuando la velocidad catalítica bordea a la velocidad de difusión (por ejemplo sustratos entrando y dejando el sitio activo, y sufriendo colisiones con otros sustratos) no representa una ventaja el aumentar aún más la velocidad de catálisis. El límite de difusión representa un cepo físico absoluto al proceso evolutivo.[1] El aumentar la velocidad catalítica más allá de la velocidad de difusión no ayuda en nada al organismo, ya que el proceso global se encuentra limitado por otro parámetro, y por lo tanto representa un máximo global en el paisaje adaptativo. Por lo tanto estas enzimas perfectas deben haber surgido de una mutación aleatoria afortunada que se extendió, o porque esta velocidad mayor alguna vez fue parte de una reacción diferente en alguno de los ancestros de la enzima.
Ejemplos
[editar]- Acetilcolina esterasa
- β-lactamasa
- Catalasa
- Anhidrasa carbónica
- Citocromo c peroxidasa
- Fumarasa
- Superóxido dismutasa
- Triosafosfato isomerasa
Véase también
[editar]Referencias
[editar]- ↑ a b c d Bar-Even, A; Noor, E; Savir, Y; Liebermeister, W; Davidi, D; Tawfik, DS; Milo, R (31 de mayo de 2011). «The moderately efficient enzyme: evolutionary and physicochemical trends shaping enzyme parameters.». Biochemistry 50 (21): 4402-10. PMID 21506553. doi:10.1021/bi2002289.
- ↑ Biochemistry, Stryer et al. 2006.
- ↑ a b c Alberty, Robert A.; Hammes, Gordon G. (1958). Journal of Physical Chemistry 62 (2): 154-9. doi:10.1021/j150560a005.
- ↑ a b c Eigen, Manfred; Hammes, Gordon G. (2006). «Elementary Steps in Enzyme Reactions (as Studied by Relaxation Spectrometry)». En Nord, F. F., ed. Advances in Enzymology and Related Areas of Molecular Biology. pp. 1-38. ISBN 978-0-470-12270-9. OCLC 777630506. doi:10.1002/9780470122709.ch1.
- ↑ a b Koenig, Seymour H.; Brown, Rodney D. (1972). «H2CO3 as Substrate for Carbonic Anhydrase in the Dehydration of HCO3−». Proceedings of the National Academy of Sciences of the United States of America 69 (9): 2422-5. Bibcode:1972PNAS...69.2422K. JSTOR 61783. PMC 426955. PMID 4627028. doi:10.1073/pnas.69.9.2422.
- ↑ Lin, Sheng-Xiang; Lapointe, Jacques (2013). «Theoretical and experimental biology in one—A symposium in honour of Professor Kuo-Chen Chou’s 50th anniversary and Professor Richard Giegé’s 40th anniversary of their scientific careers». JBiSE 6: 435-442. doi:10.4236/jbise.2013.64054.
- ↑ Chou, Kuo-Chen; Jiang, Shou-Ping (1974). «Studies on the rate of diffusion-controlled reactions of enzymes. Spatial factor and force field factor». Scientia Sinica 27 (5): 664-80. PMID 4219062.
- ↑ Chou, Kuo-Chen (1976). «The kinetics of the combination reaction between enzyme and substrate». Scientia Sinica 19 (4): 505-28. PMID 824728.
- ↑ Li, TT; Chou, KC (1976). «The quantitative relations between diffusion-controlled reaction rate and characteristic parameters in enzyme-substrate reaction systems. I. Neutral substrates». Scientia Sinica 19 (1): 117-36. PMID 1273571.
- ↑ Riggs, Arthur D.; Bourgeois, Suzanne; Cohn, Melvin (1970). «The lac represser-operatorinteraction: III. Kineticstudies». Journal of Molecular Biology 53 (3): 401-17. PMID 4924006. doi:10.1016/0022-2836(70)90074-4.
- ↑ Kirschner, Kasper; Gallego, Ernesto; Schuster, Inge; Goodall, David (1971). «Co-operative binding of nicotinamide-adenine dinucleotide to yeast glyceraldehyde-3-phosphate dehydrogenase: I. Equilibrium and temperature-jump studies at pH 8.5 and 40 °C». Journal of Molecular Biology 58 (1): 29-50. PMID 4326080. doi:10.1016/0022-2836(71)90230-0.
- ↑ Chou, Kuo Chen; Zhou, Guo Ping (1982). «Role of the protein outside active site on the diffusion-controlled reaction of enzymes». Journal of the American Chemical Society 104 (5): 1409-13. doi:10.1021/ja00369a043.
- ↑ Payens, T.A.J. (1983). «Why are enzymes so large?». Trends in Biochemical Sciences 8: 46. doi:10.1016/0968-0004(83)90382-1.
- ↑ Zhou, Guozhi; Wong, Ming-Tat; Zhou, Guo-Qiang (1983). «Diffusion-controlled reactions of enzymes: An approximate analytic solution of Chou's model». Biophysical Chemistry 18 (2): 125-32. PMID 6626685. doi:10.1016/0301-4622(83)85006-6.
- ↑ Zhou, Guo-Qiang; Zhong, Wei-Zhu (1982). «Diffusion-Controlled Reactions of Enzymes». European Journal of Biochemistry 128 (2–3): 383-7. PMID 7151785. doi:10.1111/j.1432-1033.1982.tb06976.x.
- ↑ Mireia Garcia-Viloca, Jiali Gao, Martin Karplus, Donald G. Truhlar Science 9 January 2004:Vol. 303. no. 5655, pp. 186 - 195
- ↑ Mats M. H. Olsson, Per E. M. Siegbahn, Warshel A. J Am Chem Soc. 2004 Mar 10;126(9):2820-2828.
| Control de autoridades |
|
|---|
Datos: Q408311