Can thiệp RNA
Bài viết hoặc đoạn này cần được wiki hóa để đáp ứng tiêu chuẩn quy cách định dạng và văn phong của Wikipedia. |
Bài này không có nguồn tham khảo nào. |
Trong tế bào có nhiều loại RNA khác nhau, mỗi loại đảm nhận một chức năng sinh học riêng biệt. tRNA có chức năng vận chuyển thông tin di truyền. rRNA có chức năng tham gia tổng hợp và vận chuyển protein. mRNA giúp hoàn thiện các phân tử RNA. Ngoài ra RNA còn có các chức năng quan trọng khác. Đặc biệt gần đây các nhà sinh học phân tử đã phát hiện ra chức năng điều hoà biểu hiện gene của RNA.
Kể từ khi khám phá ra RNAi thì việc nghiên cứu cơ chế và ứng dụng của nó ngày càng trở thành một vấn đề lý thú thu hút sự quan tâm của các nhà sinh học góp phần tạo nên cơn sốt "Thế giới RNA – RNA world".
Dòng thông tin di truyền
[sửa | sửa mã nguồn]Theo "Học thuyết trung tâm" (DNA RNA Protein) thì thông tin di truyền được lưu trữ ở DNA, và để tạo ra protein thì thông tin di truyền được phiên mã thành RNA và được dịch mã thành protein. Bộ gene (gene là một đoạn DNA quy định một tính trạng) của chúng ta có khoảng 30.000 gene. Tuy nhiên không phải mọi thông tin di truyền trên DNA đều dịch mã thành protein, các gene không được dịch mã gọi là non-coding gene. Các non-coding gene này tuy không được dịch mã nhưng lại tham gia vào rất nhiều quá trình trong cơ thể sinh vật như: tạo small RNA (microRNA, RNAi, hcsiRNA, piwiRNA, trans-acting RNA), được sử dụng để định danh, phân loại, tham gia vào cơ chế chống các loại bệnh, chống chịu với tác nhân môi trường,....
Khái niệm về RNAi trong tế bào
[sửa | sửa mã nguồn]"RNA can thiệp" (RNAi- RNA interference) là một loại small RNA có kích thước khoảng 19-25 nu, có khả năng tham gia can thiệp vào quá trình phiên mã và dịch mã, giúp kiểm soát các gene đang hoạt động. "RNA can thiệp" được tạo ra bên trong tế bào chất ở dạng dsDNA (double strand DNA) hình kẹp tóc và thoát ra ngoài, tiếp đó RNAi được các enzyme Dicer cắt thành dạng dsDNA hoạt tính (mature RNAi), sau đó nó được tách ra thành sợi đơn và kết hợp với AGO protein tạo thành đơn vị RISC(RNA-induced silencing complex) có khả năng ức chế quá trình phiên mã hoặc ngăn cản quá trình dịch mã. Trên động vật sẽ có các đơn vị Dicer và AGO protein khác với trên thực vật.
Dưới tác động các loại Dicer và AGO protein khác nhau sẽ tạo ra các loại RNA có tên gọi khác nhau như: microRNA, piwiRNA, RNAi, trans-acting RNA,... và chúng có chức năng khác nhau.
Vai trò RNAi
[sửa | sửa mã nguồn]RNAi có rất nhiều chức năng quan trọng trong tế bào. Chúng bảo vệ tế bào chống lại gene ký sinh trùng, virut và các yếu tố di truyền vận động (Transposon). Điều hoà biểu hiện gene. Điều khiển sự phát triển của tổ chức. Giữ gìn NST và tăng cường phiên mã. Có thể RNAi còn có nhiều chức năng khác mà con người chưa khám phá hết, và sẽ được khám phá dần trong tương lai. Các chức năng của RNAi được biết bao gồm:
- Ức chế dịch mã đơn vị mRNA.
- Ức chế sự phiên mã của gene ở trong nhân.
- Phân giải mRNA đích.
- Ức chế quá trình xâm nhiễm của vi khuẩn, nấm, virus.
- Chống chịu với các tác nhân độc hại của môi trường: tia UV, mặn, khô hạn, ngập,thiếu dinh dưỡng, kim loại nặng,...
Lịch sử nghiên cứu
[sửa | sửa mã nguồn]Trong lịch sử, sự can thiệp RNA được biết đến với những tên gọi khác như: RNA silening, quelling, cosuppresion, RNA inteference
- Năm 1984, Pesthea và các cộng sự đã nghiên cứu kỹ thuật Antiense-RNA trên vi khuẩn Escherichia Coli.
- Đến những năm đầu thập niên 1990 một số nhà khoa học công bố kết quả nghiên cứu trên các tạp chí quốc tế (Napoli và cộng sự, Vander và cộng sự).
- Năm 1994, Cogoni và cộng sự đã tiến hành thí nghiệm tăng màu cam của nấm Neurospora crassa.
- Năm 1995, trên tạp chí Cell số 81, nhóm nghiên cứu của Guo và Kemphues đã đưa ra bằng chứng đầu tiên trên tuyến trùng Caenorhabditis elegans rằng: Phân tử RNA chiều thuận (sense RNA) cũng gây ra sự ức chế gene.
- Đến năm 1998, nhóm nghiên cứu Fire đã giải thích được điều nghịch lý này bằng những thí nghiệm trên tuyến trùng C. elegans.
- Năm 2000, trên tạp chí Nature cũng công bố việc phát hiện hiện tượng RNAi trên loài ruồi giấm ProSophila do nhóm nghiên cứu của Richard Cathew tiến hành.
- Năm 2001, lần đầu tiên RNAi được mô tả trong các tế bào động vật có vú (Tuschl và cộng sự).
- Năm 2006, giải thưởng Nobel sinh lý và y học cho phát hiện cơ chế RNAi của hai nhà bác học Mỹ là Andrew Fire (ĐH Stanford) và Craig C. Mello (ĐH Massachusetts).
Khám phá cơ chế RNAi và giải thưởng Nobel y học 2006
[sửa | sửa mã nguồn]Andrew Fire và Craig Mello đã tiến hành nghiên cứu về cơ chế điều khiển biểu hiện gene ở giun tròn Caenorhabditis elegans (C.elegans). Hai ông đã thực hiện hàng loạt các thí nghiệm ngoạn mục nhằm kiểm tra kiểu hình ảnh hưởng của việc tiêm RNA vào bộ phận sinh dục của C.elegans.
Thí nghiệm nghiên cứu
[sửa | sửa mã nguồn]Tiêm các phân tử mRNA mã hoá một protein cơ bắp (sợi "có nghĩa")
=> không làm thay đổi hành vi của giun tròn.
- Khi tiêm RNA "vô nghĩa" (antisence RNA) cũng không thấy hiện tượng gì.
- Tuy nhiên, khi Fire và Mello tiêm cả RNA "sence" và "antisence" vào cơ thể giun tròn, thấy giun có những chuyển động lạ, cụ thể là co giật. Các chuyển động tương tự cũng xảy ra ở những con giun hoàn toàn thiếu một gene chịu trách nhiệm tạo protein cơ bắp.
- Khi RNA "sence" và "antisence" gặp nhau, chúng kết hợp thành RNA chuỗi đôi. Do đó, một phân tử RNA chuỗi đôi có thể làm câm lặng gene mang cùng mật mã với RNA hay không. Fire và Mello đã tiêm các phân tử RNA chuỗi đôi chứa các mật mã di truyền sản xuất nhiều protein khác nhau của giun tròn khiến gene chứa mật mã này ngừng hoạt động. Protein mã hóa gene đó cũng không được tạo ra.
- Sau các thí nghiệm thực hiện, Fire và Mello cho rằng RNA chuỗi đôi có thể làm ngưng hoạt động của các gene. Cơ chế can thiệp hữu hiệu đối với gene mà mật mã của nó khớp với mật mã phân tử RNA được tiêm.
- Cơ chế can thiệp RNA là một tiến trình xúc tác.
Những suy luận từ kết quả thí nghiệm
[sửa | sửa mã nguồn]RNA chuỗi đôi có thể làm các gene ngừng hoạt động (bất hoạt gene). Cơ chế can thiệp RNA này mang tính đặc trưng đối với gene mang mã di truyền giống với mã di truyền của phân tử RNA được tiêm vào.
Ngoài ra, cơ chế can thiệp RNA có thể lan giữa các tế bào và thậm chí được di truyền sang đời sau. Chỉ cần tiêm một lượng nhỏ phân tử RNAi cũng có thể đạt được kết quả mong muốn. Do vậy, có thể coi cơ chế can thiệp RNAi là một quá trình xúc tác.
Ý nghĩa khoa học của công trình nghiên cứu
[sửa | sửa mã nguồn]Công trình nghiên cứu này của hai nhà bác học Z.Fire và C.Mello đã được công bố trên tạp chí Nature vào ngày 19/2/1998. Kết quả của nghiên cứu này vô cùng quan trọng. Chúng cung cấp lời giải thích cho các hiện tượng nghiên cứu ở thực vật: sự bất hoạt gen sau phiên mã (PTGS- post transcriptional gene silening). Khám phá của họ đã làm sáng tỏ nhiều quan sát thí nghiệm mâu thuẫn và khó hiểu trong nhiều năm trước đây. Đồng thời nó tiết lộ một cơ chế tự nhiên để kiểm soát dòng thông tin di truyền trong tế bào, báo hiệu sự khởi đầu cho một lĩnh vực nghiên cứu mới.
RNAi được sử dụng trong khoa học cơ bản nghiên cứu chức năng của gene. Ngoài ra, cơ chế này có ý nghĩa rất quan trọng đối với việc điều khiển các biểu hiện gene, tham gia bảo vệ cơ thể chống nhiễm virus và kiểm soát gene thay đổi đột ngột. Với nghiên cứu mới này, giới khoa học cũng đang tìm ra các ứng dụng của RNAi trong những nghiên cứu y học chữa bệnh bằng liệu pháp gene, các ứng dụng trên cây trồng, vật nuôi trong nông nghiệp nhằm tạo ra các sản phẩm với chất lượng tốt hơn; trong điều trị các bệnh nhiễm khuẩn, các bệnh do virut, bệnh tim, ung thư, rối loạn nội tiết và nhiều chứng bệnh khác.
Từ kết quả của nghiên cứu này đã mở ra nhiều hướng nghiên cứu và được tạp chí Science bình chọn là "Break Through in 1998 "(Bước đột phá của năm 1998) dựa theo số lượng gia tăng cấp số nhân các bài báo khoa học đăng trên các tạp chí khoa học quốc tế hàng đầu.
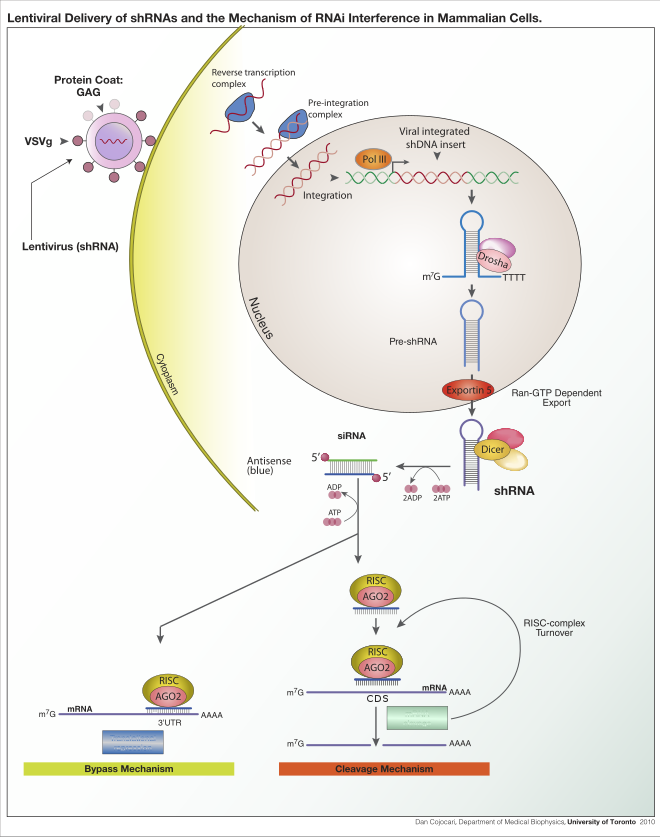
Bộ máy can thiệp RNAi
[sửa | sửa mã nguồn]Các thành phần tham gia vào quá trình can thiệp RNAi gồm siRNA và miRNA.
- siRNA
- dsRNA (double strand RNA – RNA sợi đôi) là những đoạn RNA dài mạch kép có trình tự bổ sung với gene đích (target RNA) có 2 đầu dò là 3'OH.
- Dicer: Là một loại enzyme endonuclease (Ribonuclease III) chịu trách nhiệm hoàn thiện sợi dsRNA.
- Phức hệ RISC (RNA – incluced silencing complex): Phức hệ tắt gene kích ứng bởi RNA. Phức hệ này có chứa enzyme helicase và một số protein trong đó quan trọng nhất là protein thuộc họ Agronaut (liên kết RNA) hoạt động như một endonuclea và cắt mRNA.
- siRNA (small interfeing RNA): là RNA can thiệp kích thước nhỏ được tạo ra từ dsRNA.
- miRNA là những đoạn RNA ngắn khoảng từ 19 –24 nucleotide, không tham gia vào quá trình tổng hợp protein. Bộ máy miRNA bao gồm:
- Pri- mRNA (primary- mRNA) là chuỗi mRNA nguyên thủy, dài hàng nghìn nucleotide và mang đầu 5'CAP, đuôi poly A. Pri- mRNA chứa ít nhất một hay nhiều vòng kẹp tóc (hairpin) mỗi vòng dài khoảng 70 nucleotide.
- Phức hệ RISC (RNA – incluced silencing complex): Phức hệ tắt gen kích ứng bởi RNA. Phức hệ này có chứa enzyme helicase và một số protein trong đó quan trọng nhất là protein thuộc họ Agronaut (liên kết RNA) hoạt động như một endonuclea và cắt mRNA.
- Hai enzyme cắt là Drosha ở trong nhân tế bào và Dicer ở ngoài tế bào chất
Con đường hình thành siRNA
[sửa | sửa mã nguồn]Quá trình hình thành siRNA diễn ra ở tế bào chất (cytoplasma). RNAi được kích hoạt bởi dsRNA có đầy đủ các cặp base và có ít nhất 21 -23 cặp base. Còn những phân tử dsRNA dài sẽ được là cắt thành những mảnh có độ dài khoảng 21-23 bp bởi một enzyme ngoài tế bào chất được gọi là "Dicer".
Những mảnh RNA được gọi tắt là siRNA (short interfering RNA - RNA ngắn can thiệp) và liên kết bởi protein của RNA- phức hệ gây nên sự im lặng (RISC). Những phức hệ RISC nhận ra và làm suy biến RNA sợi đơn tương ứng trong trình tự siRNA. dsRNA (double strand RNA) là đoạn RNA dài mạch kép mang trình tự bắt cặp bổ sung được với gene mục tiêu.
Khi dsRNA vào tế bào, nó bị Dicer (một loại enzyme cắt RNA mạch kép) cắt thành những đọan ngắn gọi là siRNA (small interfering RNA). Một mạch đơn của siRNA gắn với RISC (RNA - induced silencing complex) tạo phức hợp. Phức hợp này tương tác với mRNA của gene mục tiêu và cắt nhỏ mRNA này.
Trong 2 mạch đơn này, chỉ mạch nào có đầu 5' có hoạt lực với Agronauttrong phức hệ RISC mới gắn được với phức hệ RISC => tạo phức hợp siRNA-RISC. Mạch còn lại đầu 5' không có hoạt lực với Agronaut => không liên kết với RISC được. Sự xuất hiện của mạch đơn siRNA sẽ hoạt hoá RISC thành trạng thái hoạt động (RISC*).
Con đường hình thành miRNA
[sửa | sửa mã nguồn]Quá trình hình thành miRNA diễn ra ở nhân tế bào (nuclear) và trong tế bào chất (cytoplasma)
Ở trong nhân, các phân tử miRNA được tạo ra thông qua quá trình phiên mã từ các gene gọi là các phân tử miRNA nguyên thủy (pri- miRNA), các phân tử này có chứa các cấu trúc kẹp tóc (hairpin). Các phân tử pri-miRNA được cắt bởi enzyme Drosha để tạo thành những sợi Pre-miRNA (phân tử tiền microRNA)=> gọi là quá trình chế biến pri- miRNA. Các phân tử Pre- miRNA sẽ được di chuyển ra ngoài tế bào chất.
Ngoài tế bào chất, Pre- miRNA sau khi được di chuyển ra ngoài tế bào chất sẽ được enzyme Dicer cắt thành những đoạn RNA nhỏ (khoảng 19 – 21 nu). Các đoạn miRNA được tách đôi, tạo ra các sợi miRNA đơn. Trong đó sợi có đầu 5' có hoạt lực với Agronaut trong phức hợp RISC => sẽ kết hợp với phức hợp RISC tạo thành phức hệ miRNA-RISC.
Sự khác nhau giữa siRNA và miRNA
[sửa | sửa mã nguồn]Về mặt nguồn gốc, RNA mạch kép (dsRNA) có cấu trúc là chuỗi xoắn kép. Tiền miRNA(Pre-miRNA) có cấu trúc dạng thân vòng (steen-loop) hay dạng kẹp tóc (hairpin).
Xét về vị trí hình thành, chúng xảy ra ở ngoài tế bào chất. Trong nhân và ngoài tế bào chất.
Cơ chế can thiệp RNA
[sửa | sửa mã nguồn]Cơ chế chung
[sửa | sửa mã nguồn]Can thiệp RNA là một cơ chế được bảo tồn trong sinh vật đa bào nhằm giúp tế bào ức chế biểu hiện một gene khi trong tương bào có sự xuất hiện một chuỗi xoắn kép RNA có trình tự giống với gene này. Đây có lẽ là hệ thống tự vệ của tế bào nhằm chống lại sự xâm nhập của siêu vi khuẩn và các phần tử di truyền ngoại lai khác, những yếu tố sử dụng chuỗi kép RNA trong chu kỳ sống của chúng.
Khả năng chống lại các virus và gene nhảy
Sự can thiệp RNA đóng vai trò quan trọng trong việc chống lại sự xâm nhiễm của các virus, đặc biệt là các sinh vật bậc thấp hơn vào cơ thể vật chủ. Nhiều virus có thông tin di truyền chứa trên RNA sợi đôi. Khi các virus này xâm nhiễm vào tế bào, chúng bơm thông tin di truyền vào tế bào vật chủ. Khi đó, RNA lập tức vào Dicer, phức hợp RISC kích hoạt, RNA virus bị phân hủy, tế bào vật chủ thoát khỏi xâm nhiễm.
Gene nhảy (trasposon) là các trình tự DNA có thể di chuyển trong bộ gen. Nhiều transposon hoạt động bằng cách sao chép DNA thành RNA, sau đó, RNA phiên mã ngược thành DNA và gắn vào vị trí khác trên bộ gene. Sự can thiêp RNA bảo vệ bộ gene chống lại transposon.
Một số quá trình sống của tế bào có liên quan đến quá trình can thiệp RNA
- Khi virus RNA nhiễm vào tế bào, nó tiêm bộ gene có chứa RNA mạch đôi của nó vào bên trong. Can thiệp RNA tiêu hủy RNA của virus, ngăn cản sự hình thành virus mới.
- Sự tổng hợp của nhiều loại protein do các gene mã hoá cho vi RNA kiểm soát. Sau khi xử lý, vi RNA ngăn cản sự dịch mã từ mRNA thành protein.
- Trong phòng thí nghiệm, các phân tử RNA mạch đơn được biến đổi để hoạt hoá phức hợp RISC để phân hủy mRNA của một gene chuyên biệt nào đó.
- Quá trình can thiệp RNA được kích hoạt khi phân tử RNA tồn tại trong tế bào với cấu trúc sợi đôi.
- RNA sợi đôi kích hoạt một cơ chế hoá sinh để phân huỷ các phân tử mRNA có mã di truyền giống với nó.
- Khi các phân tử mRNA này biến mất => gene tương ứng bị bất hoạt => không có protein nào do gene đó mã hoá được tạo thành.
Cơ chế làm tắt gene bởi siRNA
[sửa | sửa mã nguồn]Trong tế bào, sự biểu hiện hoặc cảm ứng của RNA mạch kép dài là kết quả của sự bắt cặp giữa mạch mang mã (sợi có nghĩa) và mạch đối mã (sợi vô nghĩa). Các đoạn dsRNA sợi kép dài được cắt bởi enzyme Dicer tạo ra những đoạn RNA ngắn (siRNA) khoảng 21-28 nucleotide.
- Sau đó các siRNA được tháo xoắn dưới tác dụng của enzyme helicase và một mạch được nạp vào phức hợp protein một cách chon lọc gọi là phức hợp cảm ứng bất hoạt RISC.
- Sự xuất hiện của mạch đơn siRNA sẽ hoạt hoá RISC thành trạng thái hoạt động (ký hiệu là RISC*).
- Cuối cùng phức hợp siRNA-RISC* này sẽ tìm kiếm các transcriptome (sản phẩm của quá trình phiên mã) một cách đặc hiệu và những RNA mục tiêu tiềm năng.
- Sợi đơn siRNA sau khi được nạp vào RISC được gọi là mạch hướng dẫn, nó đóng vai trò chỉ đạo trong việc đưa phức hợp siRNA-RISC* đến các phân tử mRNA có trình tự bổ sung với nó.Việc chỉ đạo của siRNA thông qua một endonuclease có trong RISC là Agronaute protein.
- RISC* cũng có thể xâm nhập được vào nhân tế bào và kết cặp với trình tự tương đồng trên phân tử DNA hệ gene.
- Lúc này RISC* huy động một số protein làm cải biến chất nhiễm sắc quanh vị trí promoter của gene => sự kìm hãm phiên mã.
- Cơ chế tắt gen lúc này phụ thuộc vào mức độ tương đồng giữa siRNA và mRNA đích.
- Nếu sự tương đồng giữa siRNA và mRNA đích là hoàn toàn thì phân tử mRNA có xu hướng bị cắt và phân giải (do hoạt tính nuclease của RISC). => không có mRNA mã hoá cho protein đó.
- Nếu sự tương đồng giữa siRNA và mRNA chỉ là một phần thì xu hướng xảy ra là sự ức chế dịch mã do khi chung bám trên mRNA
=> ngăn cản sự dịch chuyển của Ribosome trong quá trình dịch mã => quá trình dịch mã bi ngưng lại
=> không tạo ra được protein.
- Cơ chế tắt gene bởi siRNA có hiệu quả rất cao, chỉ cần một lượng nhỏ siRNA được đưa vào tế bào cố thể đủ để làm tắt hoàn toàn sự biểu hiện của một gene nào đó (vốn có rất nhiều bản sao trong cơ thể đa bào).
Cơ chế làm tắt gene bởi miRNA
[sửa | sửa mã nguồn]- Trong tự nhiên, ngoài cơ chế điều hoà biểu hiện gene bằng sợi siRNA còn có cơ chế điều hoà biểu hiện của gene bởi một nhóm RNA khác gọi là micro RNA (miRNA).
- Cơ chế hoạt động của miRNA trong quá trình ức chế sự biểu hiện của gene cũng tương tự như ở siRNA.
- Các miRNA có chiều dài khoảng 19-24 nucleotide được tạo từ tác động cắt các đoạn Pre- miRNA có trình tự kẹp tóc (được phiên mã từ các đoạn DNA không mã hoá protein) bởi 2 enzyme là Drosha và Dicer.
- Gần 70% các miRNA được phát sinh liên quan đến sự điều tiết trong quá trình phiên mã tạo ra các mRNAvà các RNA không sinh tổng hợp các protein. Và 30% còn lại được phát sinh độc lập không liên quan đến quá trình nhân lên (chức năng của 30% miRNA này chưa được làm rõ).
- Tương tự với siRNA: Các miRNA có thể điều tiết sự phân giải mRNA với sự hiện diện của phức hợp RISC trong trường hợp bổ sung hoàn toàn(ở thực vật) (perfect complementary)
- Trong trường hợp bổ sung không hoàn toàn (inperfect complementary) với vùng 3' UTR của mRNA => ức chế quá trình dịch mã.
- Ước lượng có khoảng 120 gene mã hoá cho miRNA ở giun tròn, ở người có khoảng 250 gene và 50% gene mã hoá protein ở người đang bị kiểm soát bởi miRNA.
- Thông thường các gene miRNA được điều hoà theo kiểu chỉ biểu hiện vào những thời điểm nhất định và ở các mô nhất định trong quá trinh phát triển của cá thể.
- Đáng chú ý là 30% các phân tử miRNA ở giun trong có trình tự rất giống ở ruồi giấm và động vật có vú. Điều này cho thấy, dường như cơ chế điều hoà biểu hiện gene bởi miRNA đã có nguồn gốc từ lâu trong quá trình tiến hoá và vai trò của chúng trong việc " lập trình" biểu hiện của hệ gene là rất quan trọng đối với giới sinh vật.
Những thành tựu và triển vọng của việc nghiên cứu ứng dụng RNAi
[sửa | sửa mã nguồn]Tạo Hoa Hồng Xanh
[sửa | sửa mã nguồn]Trong cây trồng có một loại phân tử được gọi là anthocyanin được coi là sắc tố chủ đạo trên hoa, trái và các mô tế bào khác. Thông thường các màu chính của hoa bắt nguồn từ anthocyanin với sự có mặt của một ít các chất carotenoid màu vàng. Ngoài ra anthocyanin dihydrokaempferol (DHK) lại là một enzyme chi phối cho cả ba chu trình hình thành sắc tố trên cây trồng bao gồm: cyanidin, pelargonidin và delphinidin.
Gene cyanidin mã hóa một enzyme làm thay đổi enzyme DHK nhằm hình thành chu trình cyanidin dẫn đến biểu hiện các màu đỏ, hồng hay màu tím hoa cà. Trong khi đó gene delphinidin không hiện diện trong cây hoa hồng sẽ mã hóa một enzyme khá tương đồng cho việc thay đổi enzyme DHK nhằm hình thành sự tổng hợp màu theo chu trình delphinidin. Một loại enzyme khác có tên gọi là dihydroflavinol reductase (DFR) sẽ hỗ trợ các màu chỉ chị trong cả ba chu trình trên. Enzyme này rất quan trọng vì không có nó sẽ không thể tạo màu trên các cánh hoa. Chính vì vậy mà các đột biến gene DFR đều cho ra những hoa có màu trắng. Trong hoa hồng không có gene delphinidin để hình thành màu theo chu trình của nó. Chu trình delphinidin có thể hình thành màu đỏ hoặc xanh trên hoa dưới sự tác động của DRF và pH.
Để tạo ra bông hồng xanh, các nhà khoa học của Suntory đã áp dụng một bộ 3 gene. Một gene nhân tạo được dùng cho kỹ thuật RNAi nhằm ức chế gene DFR của hoa hồng làm cho hoa hồng không biểu hiện màu. Sau đó chuyển gene delphinidin từ loài hoa păng-xê và gene DFR từ loài hoa iris sẽ tạo ra hoa hồng có hàm lượng delphinidin rất cao trong cánh hoa. Tuy nhiên cũng phải lưu ý một yếu tố ảnh hưởng đến màu xanh trên cánh hoa đó chính là độ pH tế bào và đó là một trong những lý do chính là tại sao các loài hoa có cùng chu trình anthocyanin nhưng lại có màu khác nhau. Khi nồng độ pH tế bào mang tính kiềm thì sắc tố của anthocyanin thường trở nên xanh hơn. pH của đất không ảnh hưởng hay ảnh hưởng rất ít đến pH tế bào cánh hoa. Nồng độ pH tế bào cánh hoa thường mang tính di truyền. Cánh hoa hồng thông thường có nồng độ pH khoảng 4.5 chính vì vậy để tạo ra các cánh hoa hồng có nồng độ pH thấp thì rất hạn chế. Vì vậy các nhà khoa học mới nghĩ đến kỹ thuật ức chế gene bằng kỹ thuật RNAi nhằm xác định những gen ảnh hưởng đến tính axít của cánh hoa hay điều chỉnh màu của cánh hoa theo những hướng khác.
Ứng dụng cơ chế can thiệp RNA trong nghiên cứu ung thư
[sửa | sửa mã nguồn]Tế bào ung thư phát sinh từ sự tích lũy và chọn lọc nhiều đột biến có liên tiếp có lợi cho sự phân chia và tồn tại của chúng. Những biến đổi di truyền và đôi khi trên vật chất di truyền (epigenetic) giúp tế bào ung thư vượt qua sự khống chế của các nguyên tắc điều hòa tế bào và cơ thể như chương trình tự sát tế bào (apoptosis) hay các tín hiệu kìm hãm phân chia (antiproliferative signals). Sự khám phá ra cơ chế can thiệp RNA không lâu được nhận thức rằng đây chính là công cụ cần thiết để dò tìm các cơ chế phân tử bị thay đổi trong tế bào ung thư. Do tính đặc hiệu của quá trình can thiệp RNA kết hợp với tính dễ dàng nhân rộng tiến trình thực nghiệm từ hàng ngàn gene lên tới toàn bộ genome trong một thí nghiệm, hệ thống các "điểm yếu" của ung thư sẽ được giải mã và hàng loạt thuốc đặc hiệu ung thư sẽ được điều chế.
Các nhà khoa học Austaylia đã tìm ra phương pháp mang tên "con ngựa thành Troy" có khả năng tiêu diệt trực tiếp tế bào ung thư mà không gây tác dụng phụ với cơ thể người.
Cụ thể, các nhà khoa học sử dụng một tế bào nano có tên EDV để thâm nhập và làm suy yếu tế bào ung thư bằng cách tiết ra phân tử axit Ribonucleic (siRNA). SiRNA làm ngưng trệ quá trình DNA sản xuất protein vốn giúp tế bào ung thư có thể kháng thuốc hóa trị. Sau đó, một đợt EDV thứ hai sẽ tiêu diệt tế bào ung thư bằng thuốc hóa trị.
Biện pháp "con ngựa thành Troy" có khả năng tiêu diệt trực tiếp tế bào ung thư mà không ảnh hưởng tới các mô khác trong cơ thể. Hiện nay, các bác sĩ điều trị ung thư thường sử dụng phương pháp tiêm hay uống thuốc theo liệu pháp hóa trị, khiến cả tế bào lành và ung thư đều bị tiêu diệt, ảnh hưởng đến sức khỏe bệnh nhân.
Ứng dụng cơ chế can thiệp RNAi trừ sâu bệnh
[sửa | sửa mã nguồn]RNAi kiểm soát côn trùng thuộc Coleoptera. Côn trùng thuộc Coleoptera và Lepidoptera hiện được nghiên cứu về tính kháng của cây trồng nhờ protein BT sau khi thực hiện chuyển nạp gene. Một cách tiếp cận mới đối với việc kiểm soát này là sử dụng RNA can thiệp (RNAi) được các nhà khoa học của Monsanto ve Devgen N.V. thực hiện. Báo cáo khoa học được công bố trên tạp chí Nature Biotechnology.
RNAi-làm im lặng những gene cần thiết của côn trùng gây hại cây trồng, làm chúng dừng hấp thu dinh dưỡng và làm chất ấu trùng. Các nhà khoa học này đã ứng dụng RNAi để kiểm soát côn trùng gây hại rễ bắp (thuật ngữ tiếng Anh là: western corn rootworm = WCR) làm mô hình mẫu cho những nghiên cứu tiếp theo.
Phân tử RNA dây kép (dsRNA) với trình tự các cặp gốc, bổ sung cho các gen ATPase và tubulin (cytoskeletal component). Chúng thể hiện trong giống bắp biến đổi gen. Cây transgenic thể hiện sự suy giảm có ý nghĩa thiệt hại do chích hút (WCR). Tiếp cận phương pháp này, các nhà nghiên cứu còn quan sát được sự chết của ấu trùng khi phân tử dsRNA được chèn vào hai côn trùng "southern root worm" và "Colorado potato beetle". Phân tử dsRNA kiểm soát được sâu đục quả trong bông vải (boll weevil larvae), tuy nhiên, không gây ảnh hưởng đến sự chết.
Việc sử dụng RNAi để kiểm soát côn trùng gây hại cây trồng sẽ bổ sung đáng kể cho chiến lược giống chuyển gene Bt (protein diệt côn trùng) trên cây bắp, bông vải, đậu nành.
Triển vọng của việc nghiên cứu ứng dụng RNAi
[sửa | sửa mã nguồn]Việc nghiên cứu ứng dụng cơ chế can thiệp RNA có nhiều triển vọng to lớn mà con người có thể không ngờ tới được. Một số hướng nghiên cứu chính như:
1. Can thiệp RNA chống lại sự nhiễm virus.
2. Can thiệp RNA bảo đảm ổn định hệ gen bằng cách chống lại các yếu tố di truyền vận động (transposon).
3. Can thiệp RNA như cơ chế kiềm chế tổng hợp protein và điều khiển sự phát triển của tổ chức.
4. Can thiệp RNA như cơ chế giữ gìn nhiễm sắc tử cô đặc và tăng cường phiên mã.
5. Can thiệp RNA cống hiến một công cụ thí nghiệm mới để kiềm chế gene chuyên biệt.
6. Can thiệp RNA phải là một giải quyết hữu ích trong điều trị bệnh di truyền trong tương lai
Kết luận
[sửa | sửa mã nguồn]- Như chúng ta thấy, qua trên nửa thế kỷ, những nhà nghiên cứu sinh học phân tử đã thu được những khám phá ngoạn mục.
- Sự khám ra tế bào có một cơ chế đặc biệt – cơ chế can thiệp RNA (RNAi – RNA interference), đã vượt quá mong đợi và mở rộng hiểu biết của chúng ta về cơ chế kiểm soát gene trong cơ thể sinh vật.
- Việc ngày càng phát hiện ra nhiều chức năng khác nhau của RNA là cơ sở cho quan điểm cho rằng RNA có thể là đại phân tử sinh học xuất hiện đầu tiên trong quá trình tiến hoá chứ không phải là DNA hay protein như nhiều giả thiết trước đây.
- Sự phát triển của một tổ chức và chức năng chính xác của mỗi tế bào và mô tùy thuộc vào một cơ chế can thiệp RNA còn nguyên vẹn trong cơ thể.
- Hơn nữa, sự khám phá ra can thiệp RNA không chỉ cung cấp cho chúng ta một công cụ thí nghiệm mạnh mẽ mới để nghiên cứu chức năng của các gene mà còn nuôi dưỡng những mong đợi của chúng ta về những ứng dụng tương lai của can thiệp RNA trong y học, nông nghiệp,...
- Đặc biệt là Y học nghiên cứu ứng dụng RNAi để chữa những căn bệnh về tim mạch, ung thư, các bệnh vius: cúm (AH5NI, AH1N1), HIV... giải quyết tốt các vấn đề nhân loại.
Đọc thêm
[sửa | sửa mã nguồn]- Đinh Đoàn Long, Đỗ Lê Thăng; Cơ sở di truyền học phân tử và tế bào; Nhà xuất bản Đại Học Quốc gia Hà Nội; 2008
Xem thêm
[sửa | sửa mã nguồn]- RNAi: RNA inteference (RNA can thiệp)
- dsDNA: double strand DNA (DNA mạch kép dài)
- miRNA: micro RNA (tiểu RNA)
- RISC: RNA – incluced silencing complex (phức hệ tắt gene tích ứng bởi RNA)
- Dicer: RNase III (Một loại enzyme endonuclease)
- siRNA: small interfeing RNA (RNA can thiệp kích thước nhỏ)
Tham khảo
[sửa | sửa mã nguồn]Chủng loại Acid nucleic | |||||||
|---|---|---|---|---|---|---|---|
| Thành phần hợp thành |
|  | |||||
| Acid ribonucleic (biên soạn mã, không biên soạn mã) |
| ||||||
| Acid deoxyribonucleic | |||||||
| Chất tương tự | |||||||
| Vectơ tách dòng | |||||||